Description de l’habitat général de l’hirondelle de rivage
Ce document est une description technique à caractère scientifique de la région de l’habitat protégé pour l’hirondelle de rivage.
Une description de l’habitat général est un document technique fournissant une plus grande précision sur l’habitat protégé d’une espèce; cette description se fonde sur la définition d’habitat général prévue à la Loi de 2007 sur les espèces en voie de disparition.
La protection générale de l’habitat ne comprend pas une aire où l’espèce se trouvait antérieurement ou qui a le potentiel afin de l’y réintroduire, à moins que les membres existants de l’espèce ne dépendent de l’aire pour mener à bien leurs processus de vie.
La description d’habitat général décrit aussi comment la catégorie de l’habitat a été établie conformément à la « politique de catégorisation et de protection de l’habitat aux termes de la Loi de 2007 sur les espèces en voie de disparition » et est fondée sur la meilleure information scientifique disponible.
Catégorisation de l’habitat
- Catégorie 1 (rouge) : La colonie de nidification de l’hirondelle de rivage, y compris l’ensemble de terriers et le substrat entre les terriers et autour de ceux-ci.
- Catégorie 2 (orange) : La zone à moins de 50 m de la façade du talus abritant la colonie de nidification
footnote 1 pour permettre à l’hirondelle de rivage d’entrer et de sortir des terriers. - Catégorie 3 (jaune) : La zone d’habitat propice à l’alimentation à moins de 500 m de la limite externe de la colonie de nidification.
Catégorie 1
La colonie de nidification de l'hirondelle de rivage, y compris l’ensemble de terriers et le substrat entre les terriers et autour de ceux-ci, est incluse dans l’habitat de catégorie 1; on juge que celle-ci a la tolérance la plus faible à la perturbation. Les colonies de nidification sont des zones très importantes qui sont utilisées par les membres des colonies pour la reproduction, dont la ponte, l’incubation des oeufs, l’alimentation, le repos et l’élevage des oisillons.
Les trois principaux types d’habitat occupés par les hirondelles de rivage sont les falaises côtières, les berges des rivières et les sablières et gravières actives (Hickling 1959). Les colonies de nidification se retrouvent dans des emplacements discrets de talus verticaux. Les talus préférés sont verticaux (90 degrés) ou légèrement inclinés (75 degrés) jusqu’à légèrement réclinés (105 degrés) (Hjertaas 1984), bien que les hirondelles de rivage utiliseront aussi des talus avec un angle plus raide allant de 70 à 110 degrés (M. Cadman – comm. pers. 2014). Les sites avec un couvert de petits arbres et arbustes dans le haut des berges et talus sont également des emplacements de choix (Hjertaas 1984). La colonie de nidification comprend plusieurs terriers avec des chambres de nidification aménagées au bout des terriers. Les mâles se servent de leurs pattes et de leur bec pour creuser les terriers dans des parois verticales de terre ferme mais friable composée principalement de substrats silteux, sableux et loameux, qui peuvent également contenir de l’argile ou du gravier (Garrison 1999). Le taux de reproduction est meilleur dans les terriers qui sont situés plus haut dans les façades de falaise (Garrison 1999).
Les terriers sont généralement creusés jusqu’à une profondeur de 60 à 90 cm au cours d’une période d’environ cinq jours, bien que cette excavation puisse prendre jusqu’à 14 jours (Garrison 1999). Les mâles qui ne réussissent pas à attirer une femelle dans leur terrier iront creuser un nouveau terrier ailleurs pour établir un nouveau territoire (Kuhnen 1985). Il faut prendre note qu’à peu près 50 % des terriers d’une colonie de nidification sont occupés pendant une saison de nidification donnée en raison d’un surplus de terriers creusés (Garrison 1999, Cadman et Lebrun-Southcott 2013). Les terriers sont construits jusqu’à ce qu’il y ait environ 27 à 30 cm entre les terriers de la colonie (Sieber 1980 dans Jones 1987). Les hirondelles de rivage ne sont pas des oiseaux territoriaux mais les terriers et la zone qui se trouve immédiatement devant les terriers sont activement défendus (Kuhnen 1985, Garrison 1999).
Une fois qu’un couple est établi, il prolonge le bout du terrier vers le haut et sur les deux côtés pour créer la chambre de nidification où le nid est construit (Hickman 1979 dans Garrison 1999). Les nids en Ontario sont généralement plats et construits avec de l’herbe, de la paille et, parfois, des brindilles, d’autres tiges de plantes, des feuilles et des radicelles (Peck et James 1987).
On trouve certaines des plus grosses colonies de nidification avec une densité élevée d’oiseaux dans des falaises le long du rivage des lacs Érié et Ontario ainsi que sur les berges de la rivière Saugeen (COSEPAC 2013). Selon les résultats de décomptes de terriers et de sondages sur les oiseaux nicheurs, la population ontarienne d’hirondelles de rivage a été estimée à 150 000 oiseaux. La plus grosse colonie connue en Ontario comprend 3 000 couples sur la rive nord du lac Érié (Sandilands 2007). Bien qu’un couple d’hirondelles de rivage puisse établir un nid solitaire, les colonies de nidification renferment en moyenne 45 couples (Hoogland et Sherman 1976, Peck et James 1987).
Il y a une forte proportion de colonies de nidification dans le Sud de l’Ontario – dans des milieux anthropiques comme des sablières et des gravières (Peck et James 1987). Les résultats préliminaires de recherches récentes donnent à penser qu’environ 50 % des sablières et gravières du Sud de l’Ontario abritent des nids d’hirondelles de rivage (M. Cadman et M. Browning – données non publ. 2014). On ne retrouve pas autant d’hirondelles de rivage dans d’autres régions de l’Ontario et il semble que leur nombre décline rapidement à une plus haute latitude (M. Browning – comm. pers. 2014).
Des études portant sur la fidélité des oiseaux adultes au site de nidification montrent qu’entre 56 et 92 % des oiseaux adultes retournent à leurs anciennes colonies de nidification (Petersen et Mueller 1979, Szep 1990 dans COSEPAC 2013). Les hirondelles de rivage peuvent s’être adaptées à la nature instable et éphémère de leurs sites de nidification en faisant preuve de souplesse dans le choix de l’emplacement de leur colonie tout en restant fidèle à une zone générale de nidification – habituellement dans un périmètre de quelques kilomètres de l’emplacement original de leur colonie (Mead 1979). Les terriers qui sont construits dans des sols relativement stables sont parfois réutilisés, même si les hirondelles de rivage préfèrent généralement creuser de nouveaux terriers, possiblement pour éviter les risques d’infection par des ectoparasites (Turner et Rose 1989, Garrison 1999, Hopkins 2001, Alves 2008).
Une zone est requise autour de la colonie de nidification pour assurer la stabilité du talus et prévenir l’affaissement du substrat. La perturbation du sol (comme avec des excavations ou l’utilisation de machinerie lourde) à proximité de la colonie de nidification peut entraîner l’affaissement du substrat (Ghent 2001a, M. Browning – données non publ. 2014). Le danger d’affaissement dépend de l’emplacement et pourrait également être influé par divers facteurs, comme l’âge du talus, la composition du substrat, l’angle de la pente, la présence d’un surplomb et la densité des terriers. Les colonies qui sont creusées dans du sable boulant peuvent courir plus de risque de s’affaisser à la suite de pluies ou d’activités à proximité qui perturbent le sol dans la façade du talus (Ghent 2001a).
La mortalité causée par l’affaissement du substrat affecte les hirondelles de rivage, avec 3,6 % des mortalités de causes connues attribuées à l’affaissement du talus dans une étude (Mead 1979). Une autre étude a documenté l’affaissement d’environ 5 % des tunnels des hirondelles de rivage examinés (Hoogland et Sherman (1976) dans Ghent 2001a). Freer (1979) a observé que les hirondelles de rivage ne retournaient pas aux sites de nidification si la colonie produisait peu ou pas d’oisillons à la suite de l’affaissement du substrat ou de la prédation; toutefois, de nouveaux couples d’hirondelles de rivage qui n’étaient pas sur les lieux du site lors d’un affaissement peuvent coloniser le site dans des années subséquentes si le talus est encore en bon état (Freer 1979). De même, une nidification réussie dans un site augmente les chances que les hirondelles de rivage adultes retournent à ce site (Freer 1979).
Dans les habitats naturels, des mécanismes comme l’érosion et le dégagement des berges maintiennent des façades verticales qui conviennent à la nidification des hirondelles de rivage (Ghent 2001a). Dans les sites anthropiques, comme les sablières et les gravières, les hirondelles de rivage utilisent les façades verticales qui sont maintenues par les activités humaines. Sans ce maintien actif, les façades s’affaissent souvent après quelques années et ne conviennent plus à la nidification (Freer 1979, Garrison 1999, Ghent 2001b). Hickling (1959) (dans Silver et Griffin 2009) a observé que les hirondelles de rivage préfèrent une façade verticale récemment érodée pour la nidification, même si de nouveaux terriers doivent être creusés chaque année. Il est donc reconnu que dans les sites anthropiques (comme les sablières ou gravières), certaines activités qui se produisent en dehors de la saison de nidification pourraient aider à maintenir la façade des talus pour fournir un habitat convenant à la nidification, en autant que les caractéristiques importantes du talus, comme la pente, la hauteur et la composition du substrat, sont maintenues pour de futures saisons de nidification des hirondelles de rivage.
Catégorie 2
La zone à moins de 50 m de la façade du talus abritant la colonie de nidification (c.-à-d. la paroi verticale qui est directement associée à l’habitat de catégorie 1 et qui le soutient) est incluse dans la catégorie 2; on juge que cette zone a un niveau moyen de tolérance à la perturbation. Cette zone ne doit pas avoir un accès bloqué afin que les hirondelles de rivage puissent entrer et sortir des terriers sans problème.
Les hirondelles de rivage ont besoin d’un espace dégagé suffisant devant leur colonie pour pouvoir s’envoler. Dans une étude, les hirondelles de rivage de la plupart des colonies observées devaient monter moins de 1 m avant de s’envoler à 60 m de la colonie. La même étude a également trouvé que dans d’autres cas, 40 m d’espace dégagé était requis. Les sites qui exigent que les hirondelles de rivage montent plus de 1 m pour une distance de 40 m ou de 2 m pour une distance de 60 m de la façade du talus ne conviennent pas à la nidification (Hjertass 1984). La zone qui se trouve devant la façade du talus ne doit pas être bloquée pour que les hirondelles de rivage puissent disposer d’un espace horizontal de vol suffisant pour entrer et sortir des terriers sans problème.
Catégorie 3
La zone d’habitat de la catégorie 3 comprend un habitat propice à l’alimentation à moins de 500 m de la limite externe de la colonie de nidification; on juge que cette zone a un niveau élevé de tolérance à la perturbation. Les hirondelles de rivage ont besoin d’aires dégagées dans cette zone pour se nourrir d’insectes, qui constituent 99,8 % de leur alimentation (Garrison 1999). Des aires dégagées avoisinantes, comme des rivières, des lacs, des terres humides, des pâturages et des champs ouverts, sont une bonne source d’insectes volants. Des habitats terrestres dégagés comme les pâturages sont préférés pour l’alimentation tandis que les aires boisées sont évitées (COSEPAC 2013, Garrison 1999). L’alimentation pendant la saison de nidification a souvent lieu à une distance de 200 m à 500 m de la colonie (Turner 1980, Garrison 1999, COSEPAC 2013), bien que les hirondelles de rivage puissent occasionnellement se rendre plus loin pour trouver de la nourriture (Mead 1979, Andrews et Kinsman 1990).
La température joue un rôle important dans la disponibilité de la nourriture et influe sur la distance que les hirondelles de rivage couvriront pour s’alimenter. Turner (1980) a trouvé que les distances parcourues par les hirondelles de rivage au cours de la saison de nidification étaient plus grandes lorsque la température était plus fraîche (c.-à-d. 502 m ± 197 m à 16 °C ou moins), puisqu’il y a moins d’insectes disponibles lorsqu’il fait plus froid.
Activités dans l’habitat de l’hirondelle de rivage
Les activités dans la zone d’habitat général peuvent se poursuivre pourvu que la fonction de cette zone en ce qui concerne l’espèce soit maintenue et qu’on ne tue ni ne harcèle les individus de l’espèce et qu’on n’y nuise pas.
Activités généralement compatibles
- Utilisation récréative générale de sentiers actuels, comme des sentiers de randonnée à pied et en vélo
- Utilisation récréative générale des lacs, rivières et plages pour des activités comme la navigation de plaisance et la pêche
- Utilisation normale des chemins actuels
- Enlèvement ou perturbation du substrat qui ne cause pas l’instabilité du sol et/ou l’affaissement des terriers des hirondelles de rivage
- Dans les sites anthropiques, comme les sablières et les gravières, les activités (comme l’excavation) en dehors de la saison de nidification qui maintiennent la fonction de la façade du talus de la colonie de nidification en tant qu’habitat convenable pour la nidification des hirondelles de rivage pour les futures saisons de nidification (p. ex. : maintien de caractéristiques comme la pente, la hauteur et la composition du substrat)
Activités généralement non compatibles footnote 2
- Activités récréatives comme l’escalade dans des aires de l’habitat qui sont vulnérables (c.-à-d. à l’intérieur ou à proximité d’un habitat de catégorie 1)
- Enlèvement ou perturbation du substrat pendant la saison de nidification qui causera probablement l’instabilité du sol et/ou l’affaissement des terriers des hirondelles de rivage
- Activités d’aménagement qui entraînent l’enlèvement de grandes parcelles d’habitat propice à l’alimentation
- Obstruction de l’espace dans les 50 m de la façade du talus de la colonie de nidification et qui entraîne un manque d’espace de vol pour les hirondelles de rivage qui entrent et sortent des terriers
Exemple d’application de la protection de l’habitat général de l’hirondelle de rivage
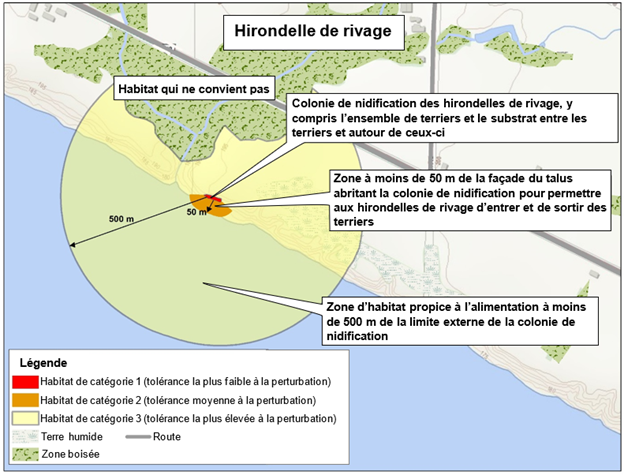
- L'habitat de l’hirondelle de rivage est situé au bord d'un cors d'eau. Une fine ligne rouge juste à l'est du cours d'eau montre un habitat de catégorie 1 (tolérance la plus faible à la perturbation). Cet habitat inclut la colonie de nidification des hirondelles de rivage, y compris l'ensemble des terriers et le substrat entre les terriers et autour de ceux-ci. Une zone orange s'étend la zone rouge jusqu'au bord du cours d'eau (50 m).
- Une zone orange s'étend la zone rouge jusqu'au bord du cours d'eau (50 m). Cette zone représente l'habitat de catégorie 2 (tolérance moyenne à la perturbation). Elle est située à moins de 50 m de la façade du talus abritant la colonie de nidification pour permettre aux hirondelles de rivage d'entrer et de sortir des terriers.
- Une zone jaune représente l'habitat de catégorie 3 (tolérance la plus élevée à la perturbation). Elle se trouve sur un rayon de 500 m autour de la zone rouge et représente la zone d'habitat propice à l'alimentation à moins de 500 m de la limite externe de la colonie de nidification. Également situés autour de la colonie de nidification sont les terres humides, une zone boisée et une route.
Réferences
Alves, M.A.S. (2008). « Effects of ectoparasites on the Sand Martin Riparia riparia nestlings ». Ibis 139 : 494-496.
Andrews, J. et D. Kinsman. (1990). Gravel pit restoration for wildlife. Royal Society for the Protection of Birds. Sandy, Bedfordshire, R.-U.
Browning, M., biologiste en recherche, ministère des Richesses naturelles de l’Ontario. Communication personnelle, le 4 juin 2014.
Cadman, M., biologiste – oiseaux chanteurs, Service canadien de la faune, Environnement Canada. Communication personnelle, le 12 novembre 2014.
Cadman, M. et M. Browning, Service canadien de la faune et ministère des Richesses naturelles et des Forêts. Données non publiées, le 11 septembre 2014.
Cadman, M. et Z. Lebrun-Southcott. (2013). « Bank Swallow colonies along the Saugeen River, 2009-2013 ». Ontario Birds 31(3) : 136-147.
COSEPAC. (2013). Évaluation et rapport de situation du COSEPAC sur l’hirondelle de rivage Riparia riparia au Canada. Ottawa. ix + 48 p. (Registre public des espèces en péril).
Freer V.M. (1979). « Factors affecting site tenacity in New York Bank Swallows ». Bird-Banding 50(4) : 349-357.
Garrison, B.A. (1999). « Bank Swallow (Riparia riparia), The Birds of North America Online » (A. Poole, éd.). Ithaca : Cornell Lab of Ornithology. (http://bna.birds.cornell.edu.cat1.lib.trentu.ca:8080/bna/species/414doi:10.2173/bna.4) [consulté le 14 mai 2014]
Ghent, A.W. (2001a). « Regular spatial patterns of Bank Swallow (Riparia riparia) tunnel entrances, with some possible evolutionary implications ». American Midland Naturalist 146 : 414-423.
Ghent, A.W. (2001b). « Importance of low talus in location of Bank Swallow (Riparia riparia) colonies ». American Midland Naturalist 146 : 447-449.
Hickling, R.A.O. (1959). « The burrow-excavation phase in the breeding cycle of the Sand Martin Riparia riparia ». Ibis 101 : 497–500.
Hickman, G.R. (1979). Nesting ecology of Bank Swallows in interior Alaska. Thèse de maîtrise. Univ. of Alaska, Fairbanks, 78 p.
Hjertaas, D.G. (1984). Colony site selection in Bank Swallows. Thèse de maîtrise. Univ. of Saskatchewan, Saskatoon, 129 p.
Hoogland, J.L. et P.W. Sherman. (1976). « Advantages and Disadvantages of Bank Swallow (Riparia riparia) Coloniality ». Ecological Monographs 46 : 33–58.
Hopkins, L. (2001). Best practice guidelines. Artificial bank creation for Sand Martins and Kingfishers. (London's BAP priority species) [consulté le 30 mai 2014].
Jones, G. 1987. « Colonization patterns in Sand Martins (Riparia riparia) ». Bird Study 34(1):20-25.
Kuhnen, K. 1985. « On pair-formation in the Sand Martin, Riparia riparia ». Journal of Ornithology 126:1-13.
Mead, C.J. 1979. « Colony fidelity and interchange in the Sand Martin ». Bird Study 26(2): 99-106.
Peck, G.K. and R.D. James. 1987. Breeding birds of Ontario: nidiology and distribution, Vol. 2: passerines. Musée royal de l’Ontario. Publications diverses sur les sciences de la vie. Toronto, 387 pp.
Petersen, C. and A.J. Mueller. 1979. « Longevity and Colony Loyalty in Bank Swallows. Bird-Banding 50(1) : 69-70.
Sandilands, A.P. (2007). « Bank Swallow », p. 394-395 dans Cadman, M.D., D.A. Sutherland, G.G. Beck, D. Lepage et A.R. Couturier, éd. Atlas of the Breeding Birds of Ontario, 2001-2005. Études d’Oiseaux Canada, Environnement Canada, Ontario Field Ornithologists, ministère des Richesses naturelles de l’Ontario et Ontario Nature, Toronto, xxii +706 p.
Sieber, O. (1980). « Causal and functional aspects of brood distribution in Sand Martins (Riparia riparia L.) ». Zeitschrift für Tierpsychologie 52 : 19-56.
Silver, M. et C.R. Griffin. (2009). « Nesting habitat characteristics of Bank Swallows and Belted Kingfishers on the Connecticut River ». Northeastern Naturalist 16(4) : 519-534.
Szep, T. (1990). « Estimation of abundance and survival rate from capture-recapture data of Sand Martin (Riparia riparia) ringing ». Ring 13 : 205-214.
Turner, A.K. (1980). The use and time and energy by aerial feeding birds. Mémoire de doctorat, University of Stirling, Stirling, Royaume-Uni. 347 p.
Turner, A.K. et C. Rose. (1989). Swallows and martins an identification guide and handbook. Houghton Mifflin Co., Boston, 258 p.
Notes en bas de page
- note de bas de page[1] Retour au paragraphe La façade du talus abritant la colonie de nidification est la paroi verticale qui est directement associée à l’habitat de catégorie 1 et qui le soutient (c.-à-d. la colonie de nidification de l’hirondelle de rivage).
- note de bas de page[2] Retour au paragraphe Si vous envisagez d’entreprendre une activité qui pourrait ne pas être compatible avec l’habitat général, s'il vous plaît contactez SAROntario@ontario.ca pour obtenir plus de renseignements.