State of the Woodland Caribou Resource Report: Part 3
The State of the Woodland Caribou Resource Report provides an overview of initiatives undertaken towards the protection and recovery of caribou. Part 3 provides a technical summary of information on the ministry’s extensive Collaborative Provincial Caribou Conservation Plan.
Cite as: Ministry of Natural Resources & Forestry (MNRF). 2014. State of the Woodland Caribou Resource Report. Species at Risk Branch, Thunder Bay, Ontario. + 156 pp.
Foreword
There are two ecotypes of woodland caribou in Ontario, which are referred to by their primary habitat: forest- dwelling woodland caribou and forest-tundra woodland caribou. The forest-dwelling boreal population of woodland caribou (Rangifer tarandus caribou) (referred to as “caribou” in this document), is listed as a threatened species under the Endangered Species Act, 2007 (ESA). As a threatened species, caribou receive both species and habitat protection. This means that harming caribou or damaging their habitat is prohibited.
The ESA requires that recovery strategies and government response statements are prepared within prescribed timelines for species listed as endangered or threatened. In 2008, the Recovery Strategy for the Woodland Caribou (Rangifer tarandus caribou) (Forest-dwelling, Boreal population) in Ontario (Ontario Woodland Caribou Recovery Team 2008) was finalized, providing scientific advice to the Minister of Natural Resources and Forestry on how to protect and recover caribou populations in Ontario. The government response statement to that recovery strategy was Ontario’s Woodland Caribou Conservation Plan (the CCP) (MNR 2009). The CCP outlines the government’s goal for the recovery of caribou. It identifies actions that the then Ministry of Natural Resources (MNR), now the Ministry of Natural Resources and Forestry (MNRF), intends to take to conserve and recover caribou in Ontario.
Ontario’s woodland caribou conservation goal
To maintain self-sustaining, genetically-connected local populations of woodland caribou (forest-dwelling boreal population) where they currently exist, improve security and connections among isolated mainland local populations, and facilitate the return of caribou to strategic areas near their current extent of occurrence.
The ESA requires a report of progress towards the protection and recovery of a species five years after publishing the government’s response statement. In addition, the CCP includes a policy commitment to developing a “State of the Woodland Caribou Resource Report” in 2014. This document meets both legislative and policy requirements.
Ontario’s Woodland Caribou Conservation Plan Progress Report (Progress Report) (MNR 2012) shares information on the accomplishments achieved in the three years since the release of the CCP. This document includes and builds on the achievements outlined in the Progress Report. While not a review or revision of the CCP, the State of the Woodland Caribou Resource Report reports on the actions the MNRF committed to in the CCP and provides a thorough overview of initiatives undertaken towards the protection and recovery of caribou. The report is divided into three parts:
Part one: Reports on MNRF’s more than 11 million dollar investment on progress made towards recovery actions and commitments in the CCP, including reporting on the status of policy, planning and resource management commitments.
Part two: Provides technical details and communicates key findings of the monitoring and assessment of caribou within Ontario’s Continuous Distribution (except Lake Superior Coast Range); describes the distribution of caribou and summarizes the findings from the initial Integrated Range Assessments.
Part three: Gives a technical summary of information on MNRF’s extensive Collaborative Provincial Caribou Research Program that discusses the findings of research commitments under the CCP.
Each part of the report can be read independently of the others but is still part of the State of the Woodland Caribou Resource Report. Additional supporting information can be found in the Appendices for each part.
Executive summary
This is the third part of the State of the Woodland Caribou Resource Report. It provides a technical summary of the implementation of a Collaborative Provincial Caribou Research Program (Research Program) as a key component of the Province’s commitment from Ontario’s Woodland Caribou Conservation Plan (CCP) (MNR 2009) to enhance caribou science.
Background: Caribou research in Ontario
Expert survey and workshops
Before the CCP’s release, the Province already recognized the importance of identifying and addressing priority caribou research needs and in the mid-2000s MNRF undertook a series of steps to develop a caribou research program for Ontario. This included an expert survey to identify key uncertainties concerning caribou ecology in Ontario (early 2006) and two expert workshops (late 2006 and 2007). The first workshop focused on translating key uncertainties about caribou into testable hypotheses. The second workshop focused on refining those hypotheses and developing an experimental framework for testing them.
Development of the research program, experimental design and other research efforts
After the second expert workshop, a group of researchers planned and implemented the Research Program to test the hypotheses that had been developed. The Research Program has been led by representatives from the MNRF, Canadian Forest Service, University of Guelph, Trent University and the Forest Ecosystem Science Co-operative, Inc. The main objective was to evaluate the degree of support for six alternative hypotheses about how human disturbance affects the long-term viability of caribou populations:
- Energetic Balance (focus: low quality food and/or increased energy expenditures)
- Sensory Disturbance (focus: human activities influences movement & habitat choice)
- Apparent Competition (focus: increased densities of alternate prey and predators)
- Predator Road Use (focus: predator road use increases hunting efficiency)
- Prey Escape (focus: fragmented habitat increases predator detection of caribou)
- Cumulative Effects (focus: combination of two or more factors are contributing to population declines)
Each of these hypotheses makes different predictions about a variety of different ecological attributes (e.g. rates of caribou energy gain and movement, predator and prey densities). To test these predictions, researchers selected three study areas, all within Ontario Shield Ecozone: one unmanaged (Pickle Lake) and two managed areas (Nakina and Cochrane). Cochrane was added in order to have a site located in the Clay-Belt portion of Ecoregion 3E (Crins et al. 2009). Study areas were chosen for their broad contrast in those characteristics believed to influence caribou persistence (e.g. conifer and mixedwood cover, linear feature densities). The primary data types for hypothesis testing included animal location and activity data, as well as high resolution video data, obtained by deploying GPS collars on woodland caribou (n=193) and wolves (n=68) within the three study areas.
The central goal of the Research Program is achieving an improved understanding of the factors that affect caribou viability. However, research efforts have also been directed towards addressing recovery actions and commitments in the CCP and testing some of the current understandings about caribou habitat in MNRF’s Forest Management Guide for Boreal Landscapes (the Boreal Landscape Guide) (OMNR 2014). At the same time as conducting the Research Program, MNRF has also been leading extensive caribou data collection efforts throughout the Continuous Distribution as part of monitoring and assessment activities (see Part 2 for more details), including the Far North Caribou Project and Integrated Range Assessments. MNRF researchers and their academic partners have also been analyzing these data to answer important questions about caribou ecotype distinctions, population delineation, status assessment methods, spatial distribution, habitat selection and space use.
General research topics
MNRF researchers have contributed to over 50 different caribou research projects that have been completed or are ongoing in Ontario. The first set of projects focuses mainly on meeting specific CCP commitments. They involve testing current understandings (e.g. in the CCP or Boreal Landscape Guide) or evaluating the effectiveness of different management approaches and explore several general topics:
- Ecotype distinction, population structure and range delineation
- Identifying best population and health measures
- Characterizing caribou habitat
- Enhancing regeneration of harvested areas
- Caribou re-occupancy of formerly harvested habitats
The second set of research projects focuses on evaluating the support for six alternative hypotheses identified by the Research Program. The third and final set of projects focuses on the development and application of models that could be used as decision support tools: namely, the caribou Population Viability Analysis (PVA) models and the Resource Selection Function (RSF) models. These models can be used to help inform assessments of population or habitat state, as well as assessments of different planning or development scenarios.
Research objectives and results
Ecotype distinction, population structure and range delineation
This research focused on clarifying the distinction between forest-dwelling and forest-tundra woodland caribou and assessing current approaches for delineating ranges in both the Far North of Ontario and southern portions of the Continuous Distribution. Results revealed clear behavioural distinctions between the forest-dwelling and forest-tundra woodland caribou ecotypes, although there was some geographic overlap in areas used by individuals from different ecotypes (particularly in winter). Caribou in the Continuous Distribution were distributed in a fairly continuous manner across the Ontario Shield and Hudson Bay Lowlands Ecozones; however, there was evidence that some degree of genetic, behavioural and demographic structure exists. This structure corresponds fairly well with the current caribou ranges that have been delineated for assessment purposes. For more information on how the ranges were delineated, please refer to the Delineation of Woodland Caribou Ranges in Ontario (Range Delineation Report) (MNRF 2014a).
Identifying best population and health measures
These research projects focused on evaluating and developing alternative approaches for assessing population state (i.e. population size estimation) and caribou occupancy patterns. Results suggested that population estimation may be feasible in some situations (e.g. for small, isolated groups), but alternative methods vary in cost and effectiveness. For situations where population size estimation is not feasible or other information is desired (e.g. spatial distribution), occupancy modelling can be a valuable tool for status assessment. The various metrics that can be used to assess population status and health have associated advantages and disadvantages. Monitoring and assessment efforts might benefit from choosing a set of population and health metrics that is effective for measuring characteristics of interest and also complementary (i.e. each compensates for the disadvantages of another metric in the set).
Characterizing caribou habitat
Several different research projects were undertaken to characterize caribou habitat at multiple scales (i.e. landscape, stand and site scales). Overall, results suggested that caribou habitat selection and time spent in an area were strongly influenced by both forage availability and predation risk avoidance, measured at fine and coarse scales. Conifer- dominated forests and treed lowlands both had year-round importance for caribou and these cover types were both selected at multiple scales. Caribou generally avoided habitats that were associated with greater risk of predation or sensory disturbance, including disturbed areas that are regenerating, deciduous and mixedwood forests, settled areas and roads. Finally, individual caribou exhibited fidelity (a tendency to return) to sites used at certain times of the year, and annual and seasonal home ranges. However, fidelity was found to be strongest for calving sites and home ranges used during the calving and post-calving seasons. Fidelity was found to be weakest for winter home ranges.
Enhancing regeneration of harvested areas
Several different research projects were undertaken to inform efforts to promote the regeneration of caribou habitat after forest harvesting. Researchers focused on a) determining how forest composition and structure changes in response to disturbance type and age, and b) evaluating the effects of stand-level silvicultural (tree care) treatments on lichen (a major caribou food item). Results indicated that plant community composition differed between natural and harvest-origin stands and that the degree of difference increased with stand age. Differences in forest structure were not as pronounced, but canopy closure was higher in older harvest-origin stands. Lichen abundance was influenced by several stand characteristics. Higher abundances were associated with the following:
- conifer-dominated cover
- low canopy closure
- stand age (i.e. older)
- non-organic sandy soils
- relatively low stand density, tree and crown height
Research on the short and long-term effects of applying different silvicultural treatments indicated that herbicide applications had negative short-term and long-term effects on lichen abundance (although impact strength varied by type). One herbicide treatment (2, 4-D) was associated with markedly different lichen composition from all other silvicultural treatments and natural-origin stands.
Caribou re-occupancy or use of formerly harvested habitats
To improve our understanding of the longer-term impacts of forest management activities on caribou habitat use, researchers are trying to identify caribou re-occupancy of previously harvested areas and characterize the attributes associated with such areas. Preliminary results from an analysis of previously harvested areas that are used by caribou during summer suggest that these areas tended to be conifer- dominated, younger than other available previously harvested areas and with different structural characteristics. Use of these areas is infrequent and more likely to be latent rather than an indication of re-occupancy; in other words, caribou may be continuing to use a recently-harvested, former use area rather than returning to or adopting an area that has sufficiently regenerated to a level that can provide suitable habitat. Therefore, while these examples may provide some insight into what might underlie the relatively infrequent summertime use of recently harvested areas by caribou; these preliminary results are not appropriate for identifying characteristics associated with longer-term re-occupancy or use of suitable caribou habitat. Additionally, evidence from other studies suggests that use of recently harvested areas may place caribou at higher risk of predation.
The energetic balance hypothesis: Diet, nutrition, energy expenditure and condition
To evaluate the level of support for the Energetic Balance Hypothesis, researchers quantified several different aspects of caribou energetics, including energy intake (through eating) and expenditures. There have also been preliminary efforts to determine whether caribou condition differs between managed and unmanaged landscapes. Results for energy intake indicate that lichens (particularly ground species) were the dominant year-round component of caribou diets, but they also had relatively low nutritional value. In terms of energy expenditure, local environmental conditions (e.g. forage availability, snow depth and temperature) and daily movement rates had a strong influence on caribou energy costs. Insect harassment was worse in open harvested areas than treed stands, but preliminary results indicated that energetic consequences for caribou may relate more to lost feeding opportunities rather than increased energy expenditure associated with efforts to avoid insects. Calf and adult condition data were limited, but preliminary evidence suggests caribou may have experienced negative nutritional impacts in some managed areas.
The sensory disturbance hypothesis
To assess the degree of support for the Sensory Disturbance Hypothesis, researchers characterized seasonal variation in traffic volume in managed and unmanaged areas and studied the effects on caribou behaviour of vehicle traffic on a major road. Traffic volume varied between seasons and study areas (lower in unmanaged areas). Preliminary results for caribou proximity to the road and road crossing rates suggested that caribou avoidance of roads may have increased (in a nonlinear manner) as traffic volume increased. Additional research focused on evaluating the effectiveness of road decommissioning and reclamation measures. Preliminary results indicate that different decommissioning strategies varied in effectiveness. In addition, if vegetation regeneration on old roadbeds is an objective, research indicated that road rehabilitation efforts will likely benefit from reclamation efforts.
The apparent competition hypothesis
To evaluate the level of support for the Apparent Competition Hypothesis, research was directed towards testing the various predictions the hypothesis makes: niche separation between caribou, moose and their shared predators; niche overlap between moose and wolves; higher densities of moose and wolves in managed landscapes; and dominance of moose in predator diets. Results from research into the occupancy, movements, habitat selection and activity patterns of caribou, moose and wolves indicated that there was a high level of overlap between wolves and moose and a high degree of spatial and temporal segregation between caribou and both of these species. Moose and wolf densities were higher in managed landscapes and wolf territories were smaller. Wolf diet analyses indicated that moose were the primary year-round prey of wolves and these predators selected landscape features that were often associated with higher moose density (i.e. mixedwood, deciduous or regenerating forests). Caribou appeared to be only the secondary or tertiary prey items for wolves throughout the year. The relationship between bears and caribou was not well studied, but analysis of bear diets during calving season yielded no appreciable evidence of caribou.
The predator road use hypothesis
To evaluate the degree of support for the Predator Road Use Hypothesis, researchers studied wolf habitat selection. They also examined the factors that influenced time between wolf kills, as well as attributes associated with kill sites of moose (their dominant prey). Results from habitat selection analyses indicated that wolves selected resource access roads at both the pack territory and broader landscape scales. Proximity to roads was also a major determinant of how efficient wolves were at hunting moose, and how likely it was that they were able to make a successful moose kill.
The prey escape hypothesis
To assess the level of support for the Prey Escape Hypothesis, research efforts focused on testing its two major predictions for managed landscapes: restricted space use among caribou in response to lower abundance and/or fragmentation of suitable habitat, and increased targeting of caribou by wolves in managed landscapes. Results from the Far North of Ontario indicate that caribou home ranges were larger in areas with low amounts of preferred habitat and smaller when the amount of suitable caribou habitat was high. Results from more heavily managed regions indicate that fidelity (the tendency to return) of individual caribou to seasonal ranges was generally high, and in most cases was not influenced by associated habitat conditions. One exception was fidelity to winter ranges. When the winter ranges of individual caribou were located close to roads and young forests, they were more likely to return to those same winter areas from year to year. In contrast, the tendency of individuals to return to the same winter ranges was lower when they were close to more suitable habitat. However, none of the findings from analyses of wolf diet, habitat selection, or the attributes associated with hunting success indicated that wolves were targeting caribou in managed landscapes.
Caribou mortality factors: Evaluating support for multiple alternative hypotheses
All of the alternative hypotheses make predictions about caribou mortality rates, mortality causes and the attributes associated with mortality events. Data on adult caribou deaths and parasite infection levels have allowed researchers to test these predictions and to address CCP commitments to increase understanding of human-caused mortalities and parasite impacts on caribou. Preliminary results suggest that wolf predation was the dominant cause of adult caribou deaths and no human-caused mortalities were documented. Bear predation was documented but was fairly uncommon. While annual survival rates were relatively high (>75%) they were lower in the most intensively managed study area (Nakina) and this difference appeared to be driven by higher predation-related mortality rates. Mortality risk varied seasonally, but was highest from late fall to early spring.
Data on the fates of caribou calves were limited to video evidence from the small subset of cows that were fitted with GPS-collars that contained video cameras. There was little evidence of mortality causes amongst calves, but preliminary results suggest most calves in managed and unmanaged areas died during the first two months after birth (i.e. from mid- late May to early July). Preliminary results from research into parasite infection levels among adult caribou indicated that incidence of Protostrongylid parasite infections may be higher in managed vs. unmanaged study areas, but there was no confirmed evidence of brainworm (i.e. P. tenuis) infections.
Caribou population viability analysis models
One of the principal goals of the Research Program was to develop Population Viability Analysis (PVA) models, which can help users assess the long-term probability of caribou population persistence and allow them to assess different potential development and management activities (e.g. new roads, changes in moose or wolf harvest rates) on caribou population viability. Two models were developed: a simple, single species PVA model that incorporates data on key vital rates (i.e. survival and recruitment estimates); and a spatially explicit PVA model that includes a caribou movement model and incorporates the major relationships (e.g. among caribou, moose and wolves) and habitat associations identified through the Research Program. Preliminary results from efforts to apply both models to populations in an unmanaged study area (Pickle Lake) and a managed study area (Nakina) suggest that the population in the managed study area may have a relatively low probability of persisting over the long-term under current conditions.
Caribou resource selection functions
Researchers developed models of caribou habitat selection at the broader landscape scale using Resource Selection Functions (RSFs). These models can be used to map suitable caribou habitat and are contributing to efforts to integrate caribou habitat considerations into planning initiatives in the Far North of Ontario. RSF results indicated that caribou habitat selection was strongest at the 10,000 ha scale and was primarily influenced by predator avoidance. Results also indicated that multiple RSF models are required for range-wide assessment of caribou habitat, as habitat selection patterns differed among regions, ranges (within regions) and seasons.
Synthesis: Implications of research results
Support for alternative hypotheses
The six alternative hypotheses evaluated as part of the Research Program were associated with variable degrees of supporting evidence. Specifically, among the single mechanism hypotheses, there appeared to be a high degree of support for Apparent Competition, a moderate level of support for Predator Road Use, Sensory Disturbance, and Energetic Balance and a low level of support for Prey Escape. It appeared likely that more than one set of factors may be contributing to caribou population declines in managed landscapes. Therefore, it can be argued that the Cumulative Effects Hypothesis had the greatest level of support from research results.
Evidence from recent research efforts also provided strong support for many pre-existing understandings about caribou biology and ecology. Weaker or mixed support was found for the following expectations: nutrition is not limiting for caribou; caribou habitat selection is driven by different factors at different scales; caribou show strong individual fidelity to seasonal home ranges used during calving (strong support) and winter (weaker support).
A synthesis of research findings from multiple projects yielded a variety of different insights:
- Caribou habitat
- Habitat selection patterns appear consistent with the influence of forage availability and predation risk avoidance at multiple spatial scales (vs. a hierarchy of influence at different scales).
- Year-round dominance of ground lichen in caribou diets was unexpected (particularly in summer) and suggests the year-round importance of conifer-dominated stands for foraging.
- General caribou-habitat relationships, such as selection of conifer-dominated forests and treed lowlands and avoidance of disturbed, settled and open areas and roads, were found to be consistent with current management approaches.
- Predation risk in managed landscapes
- Higher predator densities and greater amounts of alternate prey and predator habitat in managed landscapes can increase predation risk for caribou.
- Harvesting and silvicultural practices that promote regeneration of caribou habitat in amounts and arrangements similar to what occurs naturally would likely help ameliorate some of harvesting’s potential impacts on caribou.
- Dumpsites can provide major food subsidies for wolves and may support higher predator densities. This may result in increased predation risk for caribou.
- The influence of roads on caribou is complex; resource access roads are associated with higher risk of predation by wolves, but the dominant impacts associated with roads with high volumes of vehicle traffic may bemore related to caribou avoidance of traffic-related Efforts to mitigate the effect on caribou of roads might benefit from selecting approaches that address the dominant impacts associated with different road types.
- Habitat conditions in managed landscapes do not appear to have led to improved detection and active targeting of caribou by wolves; however, the size and connectivity of areas of suitable habitat might impact caribou in other ways (e.g. increased exposure to predation risk when moving between suitable areas).
- Harvesting and silvicultural practices that promote regeneration of caribou habitat in amounts and arrangements similar to what occurs naturally may help mitigate the impacts of resource management activities on caribou, but such approaches may be insufficient without concurrent efforts to limit the cumulative amount of disturbance in caribou ranges to levels likely to support self-sustaining populations.
- Reduced physical condition in managed landscapes (direct and indirect effects)
- There was moderate support for two hypotheses that predict reduced physical condition in managed landscapes (i.e. Energetic Balance and Sensory Disturbance) and preliminary results indicate that caribou in managed landscapes might be in poorer physical condition.
- Applying silvicultural treatments that promote ground lichen regeneration within suitable sites (i.e. those with non-organic, sandy soils) might increase caribou forage availability at the stand level.
- Harvesting and silvicultural practices that promote regeneration of caribou habitat in amounts and arrangements that are similar to patterns likely to be generated by natural disturbance regimes might increase landscape-scale forage availability (and, possibly, reduce the energetic costs of acquiring food).
- Re-occupancy of harvested stands by caribou
- Most examples of caribou use of stands that were previously harvested documented through research conducted to date, likely represent latent use of harvested areas ( vs. long-term re-occupancy or use by caribou of areas that have regenerated into suitable habitat). As such, evidence of stand characteristics associated with this use is likely not an appropriate source of information for efforts to improve silvicultural practices.
- The PVA and RSF models: Decision support tools for assessment & scenario analysis
- Extensive research efforts throughout Ontario have enabled the development of quantitative models (i.e. the PVAs and the RSFs) that incorporate key aspects of caribou ecology and behaviour. These models can be used to help assess population and habitat state. They can also be applied to landscapes that represent alternative management, development and climate change scenarios and be used to evaluate different scenarios on the likelihood that caribou will persist over the long-term and on the amount and arrangement of suitable caribou habitat.
Knowledge gaps and future research efforts
There are still some major knowledge gaps regarding caribou ecology and behaviour in Ontario. We have yet to collect direct information on causes of caribou calf deaths (including the potential role of black bears as predators), comprehensive information on non-predation mortality causes among adult caribou and fuller information on the re-occupancy of previously harvested stands. Future research efforts could be directed towards addressing these gaps, although completing such research may be challenging. A second option for future research involves expanding completed or ongoing studies to additional areas to improve replication and decrease uncertainty about the representativeness of results collected to date. A third option involves focusing on the key hypotheses supported by existing research and testing the effectiveness of different management strategies for reducing the impacts of their associated mechanisms (e.g. apparent competition, predator road use) on caribou.
1 State of the Woodland Caribou Resource Report: Part 1
The State of the Woodland Caribou Resource Report provides an overview of initiatives undertaken towards the protection and recovery of caribou. Part 1 reports on the progress of the recovery actions and commitments in the Caribou Conservation Plan, which has been supported by a more than $11-million investment, as well as the status of policy, planning and resource management commitments.
2 State of the Woodland Caribou Resource Report: Part 2
The State of the Woodland Caribou Resource Report: Part 2 provides technical details and outlines key findings of the monitoring and assessment of caribou within Ontario’s Continuous Distribution (except the Lake Superior Coast Range); describes the distribution of caribou and summarizes the findings from the initial Integrated Range Assessments.
3 Collaborative Provincial Caribou Research Program
3.1 Background information
The importance of identifying key caribou research needs and of undertaking research that addresses significant conservation and management concerns in Ontario has been recognized since the end of the 1990s (e.g. “Planning the next Round of Caribou Habitat Research”, Apr. 29-30, 1999 – Brown 1999). This awareness gave rise to several formal MNRF-led efforts to identify high priority research questions for caribou conservation in the province, and to develop a collaborative caribou research program to help answer them.
In early 2006, an expert survey was distributed to MNRF managers and staff, resource industries, non-governmental organizations, academics, tourism representatives, and federal and municipal government representatives known to have had involvement or interest in caribou (Rodgers et al. 2007). Participants were asked to rate the importance of several general research subject areas and specific research questions. Key uncertainties identified by survey respondents were all related to the direct and indirect effects of disturbance (e.g. by commercial forestry operations, mining, or fire) on caribou in Ontario. Specific issues identified as priorities for future research included (Rodgers et al. 2007):
- Habitat selection by caribou at multiple scales;
- Impacts of predators; and
- Impacts of roads and other linear features.
Following this survey, MNRF organized two expert workshops that were focused on developing a caribou research program for Ontario. The first, held at the end of 2006, included participants from the provincial governments of Ontario, Alberta, Manitoba and Québec, academics, and representatives from several different non-governmental organizations and forestry companies (Rodgers et al. 2007). The objective of this workshop was to review the issues identified in the survey and refine them into hypotheses that could be evaluated through scientific research. Alternative hypotheses developed at the workshop focused on the impacts that habitat pattern, roads and predation have on the probability of caribou occurrence. The general working hypothesis that emerged from this first workshop was that the probability of caribou occurrence is a function of roads (density/use/type), predation (predator density/predation rate/prey biomass or abundance) and multi-scale habitat pattern.
The second workshop, held in 2007, involved a more focused group of participants with expertise in scientific research and experimental design, and included researchers from the Ontario, Manitoba and Québec governments, the federal government and various academic institutions (Rodgers et al. 2008). The objective of this second workshop was to review the hypotheses developed in the first workshop and develop a framework for a set of studies and experiments that would allow researchers to test these hypotheses. Participants concluded first, that estimating the probability of caribou persistence ( vs. probability of caribou occurrence) was the most appropriate way to address the research questions identified in the first workshop and second, that developing population viability models and conducting population viability analyses was the most appropriate framework for testing many of these hypotheses.
Workshop participants also identified the different types of study areas that would need to be used and the different dependent and independent variables that would need to be measured to assess evidence for alternative hypotheses about the factors that affect caribou persistence. Subsequent to the workshop, a subset of potential study areas was identified (Rodgers et al. 2009).
Following the second workshop, a group of researchers came together to plan and implement a Collaborative Provincial Caribou Research Program (Research Program) to test the hypotheses outlined in the second workshop. The program was led by a team of researchers with representatives from the MNRF, Canadian Forest Service (CFS), University of Guelph, Trent University and the Forest Ecosystem Science Co-operative, Inc.
The many studies undertaken as part of this Research Program have been influenced by the findings and recommendations of the two expert workshops and by the key uncertainties and research priorities regarding caribou identified in the CCP (MNR 2009). The implementation of the Research Program and the consideration of research results when developing caribou conservation and management approaches are two of the key action items listed in the CCP (MNR 2009).
Studies completed or underway are summarized in Section 3.2. Summaries focus on study objectives, methods used, results to date and remaining knowledge gaps. Direct links between the research described and the different action items that the Government of Ontario committed to in the CCP (MNR 2009), are also indicated. The results from the research results provide insights to caribou ecology and biology which are discussed in detail in Section 3.3.
3.2 Summary of progress and results from the Collaborative Provincial Caribou Research Program
3.2.1 General overview of research projects and report structure
Research program design and data collection
The main focus of the Research Program has been to evaluate the degree of support for several alternative hypotheses about the key factors affecting caribou population persistence, each of which focuses on contrasts between managed (i.e. human disturbed) and unmanaged forests. Alternative hypotheses include:
- Energetic balance – Managed forests have lower quality & quantity of caribou food and/or are associated with increased energetic costs of obtaining food.
- Apparent competition – Managed forests attract and support high densities of moose and deer, which results in increased densities of wolves and bears and increased predation rates on caribou.
- Predator road use – Roads developed in managed forests are used by predators, leading to increased hunting efficiency and higher predation rates on caribou.
- Prey escape – In managed forests, caribou are restricted to fewer and smaller patches of mature conifer forest, which increases search effectiveness of predators.
- Sensory disturbance – Noise and human activity in managed forests influence caribou movement and/or habitat choice.
- Cumulative effects – Some combination of two or more of the preceding hypotheses contributes to the decline of caribou populations.
Each hypothesis is associated with a different set of predictions regarding several different attributes related to the ecology of caribou in the different study areas; including rates of caribou energy gain, predation rates, moose and deer densities, caribou movement rates and caribou survival and reproductive rates. Comparing patterns in the data collected to these predictions enables researchers to determine which factors have the greatest influence on caribou decline, which in turn will allow them to develop recommendations for improving caribou conservation and management efforts.
To test these predictions and determine the degree of support that exists for the alternative hypotheses outlined above, the Research Program identified nine candidate study areas within the Continuous Distribution (Rodgers et al. 2009). These candidate areas were distributed from east to west along the northern extent of forestry operations within the province and were identified based on recommendations developed at the experimental design workshop (Rodgers et al. 2007). Candidate study areas were assessed for suitability by evaluating the state of key attributes that are believed to affect the probability of caribou persistence, including conifer and mixedwood cover, natural and anthropogenic disturbance, as well as predator, alternate prey, and linear feature densities (Rodgers et al. 2009).
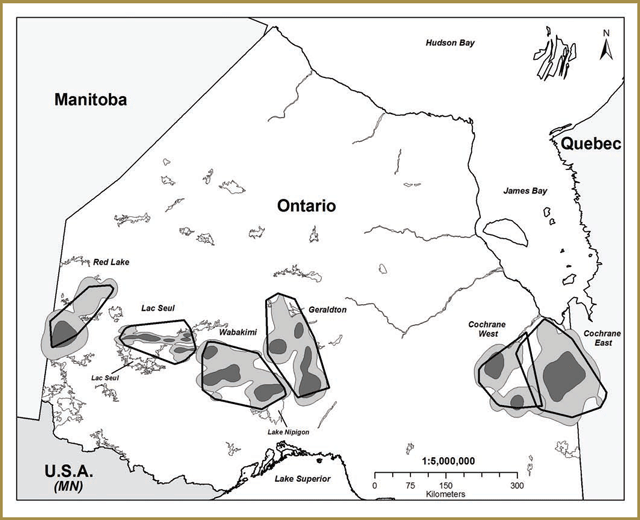
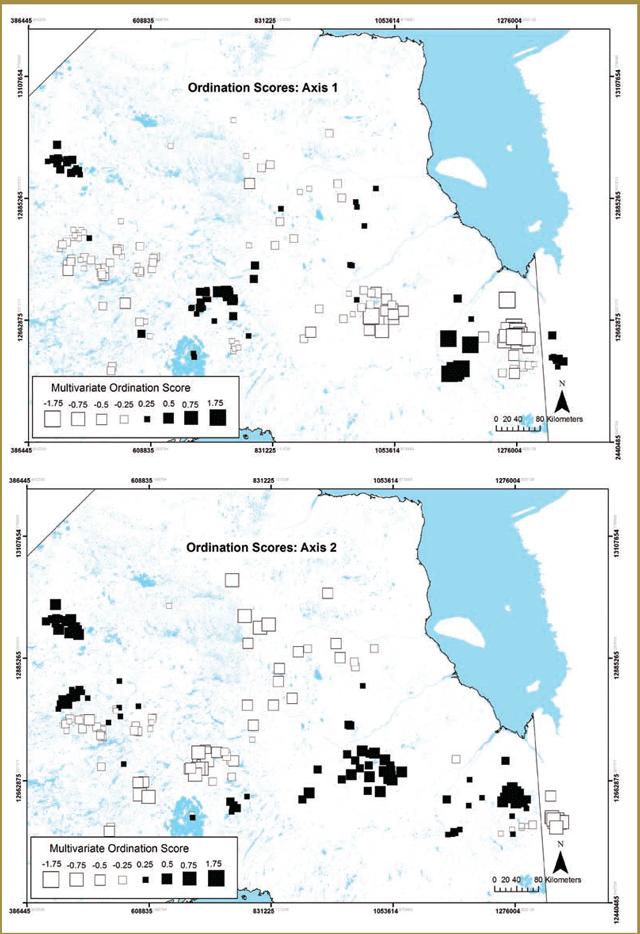
Researchers initially chose two of these study areas for focused data collection based on efforts to obtain broad contrasts with respect to the attributes listed above: Pickle Lake (unmanaged forest) and Nakina (managed forest). Pickle Lake corresponds broadly with the Kinloch Range and Nakina generally overlaps with the Nipigon Range. A third study area, Cochrane (managed forest), which generally corresponds to the Kesagami Range, was added a year later. All three study areas are located within the Ontario Shield Ecozone (Figure 3-1), with the Cochrane study area being located within the Clay-Belt portion of Ecoregion 3E (Crins et al. 2009).
Pickle Lake has high conifer cover, low wolf and moose densities and the lowest level of anthropogenic disturbance (i.e. linear feature development and forest harvesting) (Figure 3-1). Nakina has the lowest conifer and highest mixedwood cover, high wolf and moose densities and the highest level of anthropogenic disturbance (i.e. linear feature development and forest harvesting) (Figure 3-1). Cochrane has the highest conifer and lowest mixedwood cover, low wolf densities, low moose densities and intermediate levels of human disturbance (Figure 3-1).
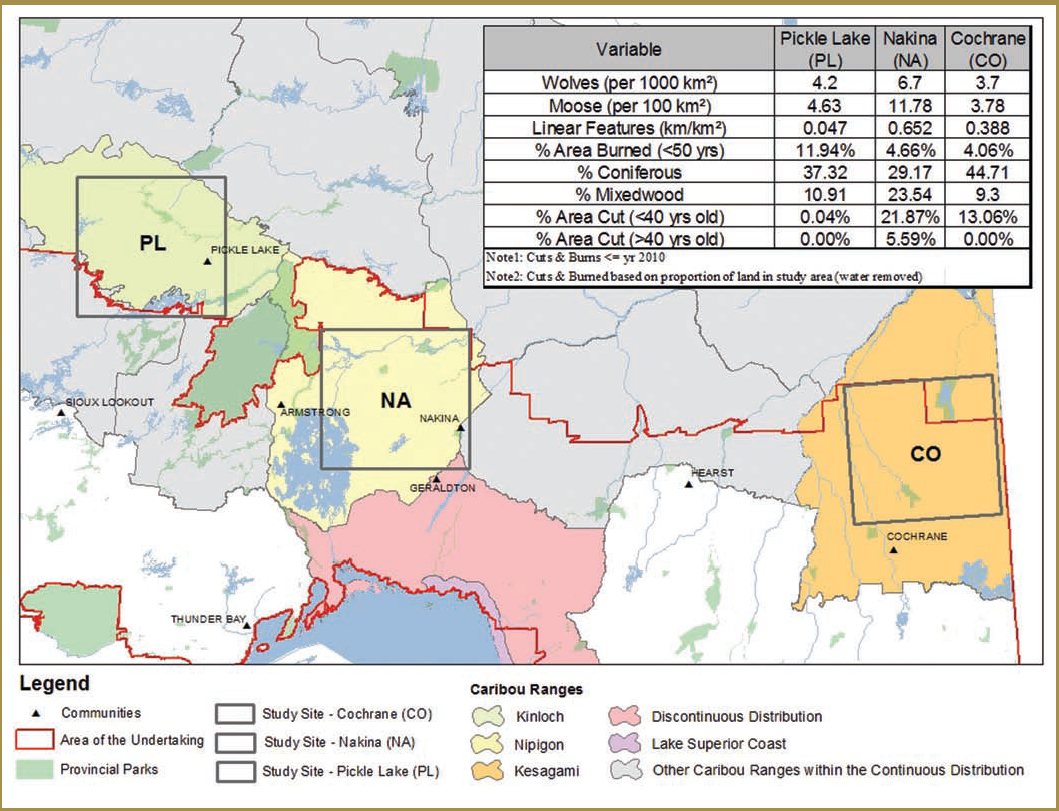
| Variable | Pickle Lake (PL) | Nakina (NA) | Cochrane (CO) |
|---|---|---|---|
| Wolves (per 1000 km2) | 4.2 | 6.7 | 3.7 |
| Moose (per 1000 km2) | 4.63 | 77.78 | 3.78 |
| Linear Features (km/km2) | 0.047 | 0.652 | 0.388 |
| % Area burned (<50 years) | 11.94% | 4.66% | 4.06% |
| % Coniferous | 37.32% | 29.17 | 44.71 |
| % Mixedwood | 10.91 | 23.54 | 9.3 |
| % Area cut (<40 years old) | 0.04% | 21.87% | 13.06% |
| % Area cut (>40 years old) | 0.00% | 5.59% | 0.00% |
Note 1: Cuts &burns <=year 2010
Note 2: Cuts & burned based on proportion of land in study area (water removed)
The primary data source for the Research Program has been animal location and activity data, as well as high resolution video data, all of which were obtained by deploying GPS-collars on caribou and wolves within the three study areas. In each of the study areas a small subset of collared caribou were fitted with specialized GPS-collars with high resolution video cameras. Collaring efforts began in the winter of 2010 and continued every winter until 2013.
Caribou collaring efforts were guided by several objectives:
- To maintain functioning collars on at least 50 adult female caribou per study area for 3 consecutive years;
- To ensure collars were deployed in study areas large enough to support a viable caribou population of approximately 200–400 individuals (i.e. at least 5,000 – 10,000 km2; Rodgers et al. 2008); and
- To deploy collars in a manner that was representative of the observed distribution of caribou throughout the study area.
Wolf collaring efforts were guided by similar objectives:
- To maintain functioning GPS-collars on at least one adult wolf and VHF collars on at least two adult wolves per pack in each study area for 3 consecutive years.
- To deploy collars in a manner that was representative of the observed distribution of wolves throughout each study areas.
Data from a total of 193 GPS-collared caribou and 68 GPS-collared wolves have been and continue to be analyzed as part of the Research Program. Although black bears were identified as a major potential predator of caribou (particularly of calves) during the project development process (Rodgers et al. 2006, Rodgers et al. 2008), the considerable resources required to capture and collar bears were not available, therefore, no bear telemetry data were collected as part of the project.
While improving understandings of the ecological processes and interactions that influence population persistence amongst caribou is the central goal of the Research Program, additional research efforts have also been directed towards addressing some of the recovery actions and commitments made in the CCP (MNR 2009). These include examining local population structure and exploring alternative approaches for determining population status. Research efforts have also been directed towards testing some of the current understandings of caribou habitat that are included in MNRF’s Forest Management Guide for Boreal Landscapes (‘Boreal Landscape Guide’) (OMNR 2014), determining what factors influence caribou re-occupancy of harvested areas (CCP 1.2), evaluating the effectiveness of different silvicultural approaches for enhancing caribou habitat regeneration (CCP 4.1.2), and the effectiveness of different road decommissioning and rehabilitation techniques (CCP 4.2.1).
Other research efforts
In addition to the data collected for the Research Program, MNRF has been leading extensive caribou data collection efforts throughout the Continuous Distribution since 2009 through monitoring and assessment activities (see Part 2 for more detailed information) such as the Far North Caribou Project and Integrated Range Assessments. Data collected as part of the monitoring and assessment activities includes observation data from systematic aerial surveys (see Part 2, Section 2.3.1) and telemetry data from GPS-collars deployed across the province (see Part 2, Section 2.4). In addition to monitoring-related uses, MNRF including researchers from Ontario Parks and their academic partners have also been analyzing these data to answer some important questions about ecotype distinctions, population delineation, status assessment methods, caribou behaviour and spatial distribution. Descriptions of these research initiatives are also included in Part 2. Finally, in addition to research that is focused (wholly or in part) on forest-dwelling woodland caribou, the MNRF has also been a partner in recent research that focuses on forest-tundra woodland caribou (e.g. Newton 2012, Newton et al. 2014, Newton et al. in review). While a detailed description of research that focuses on the forest-tundra ecotype is beyond the scope of this report, it explores topics like the recent change in calving grounds used by the Pen Islands herd, the impacts that caribou can have on their traditional areas and the consequences of those effects for long-term patterns of population distribution. Consequently, although not described in further detail in this report, this research also has led to an improved understanding of the ecology and biology of both caribou ecotypes.
Overview of the report: structure and content
The rest of Section 3.2 (i.e. 3.2.2 - 3.2.14) focuses on describing the diverse caribou research efforts that MNRF has been involved with in recent years. General goals and objectives, methods and findings and the implications of research results for caribou conservation and recovery are also discussed. All research projects are grouped by general topic and the projects that are directly related to the CCP and Boreal Landscape Guide are described first: Population Structure & Range Delineation; Identifying Best Population and Health Measures; Characterizing Caribou Habitat; Enhancing Regeneration of Harvested Areas; and Caribou Re-occupancy of Formerly Harvested Habitats. Research that has strong implications for the five single factor hypotheses examined by the Research Program are described second: Energetic Balance; Sensory Disturbance; Apparent Competition; Predator Road Use, Prey Escape; and Caribou Mortality Factors. Finally, two projects that focus on synthesizing multiple research results to explore the potential implications of different management and development actions on caribou population persistence or probability of caribou use are also described: the Population Viability Analysis and the Resource Selection Functions (RSFs).
The bracketed numbers that appear in Sections 3.2.2 to 3.2.14 (e.g. [Project # 23. Please see Appendix 3-1]) represent unique research project numbers that can be used to find relevant project information in Appendices 3-1 and 3-2. Appendix 3-1 provides a general overview of all research efforts described in Section 3.2 and includes information on lead researchers and organizations involved in specific research projects, along with study locations, the hypotheses being evaluated and the action items in the CCP that research results contribute to. Appendix 3-2 contains a list of publications associated with different research projects (i.e. articles in peer-reviewed journals, graduate student theses, research manuscripts submitted to peer-reviewed journals for review and government reports). All research findings that have not been published as a peer-reviewed journal article, a defended and completed graduate student thesis, or as a portion of a government report are described as preliminary results.
The closing section of Part 3 (Section 3.3) summarizes and synthesizes key findings from recent caribou research efforts in Ontario. It also includes general discussion of the overall implications of research results for caribou ecology and biology. Topics examined include caribou habitat, predation risk and reduced condition in managed landscapes, re-occupancy of previously harvested areas, as well as the application of different scenario analysis approaches that can be used to help inform assessments of potential impacts to caribou and their habitat. Finally, remaining gaps in caribou knowledge and future research directions are discussed.
3.2.2 Ecotype distinction, population structure and range delineation
Researchers found behavioural differences between the forest-dwelling and forest-tundra woodland caribou ecotypes, even though the landscapes they used overlap during the winter.
Research indicates that, although caribou were continuously distributed across the Ontario Shield and Hudson Bay Lowlands Ecozones, they also demonstrated some degree of genetic, behavioural and demographic differences throughout their distribution.
Research results suggest that the current boundaries for the southern caribou ranges were generally supported by evidence from both historic and recently collected data on caribou movement.
Research goals & objectives
The CCP outlines an approach to the management and recovery of caribou that includes applying a Range Management Approach to conserve and recover the ecotype throughout their Continuous Distribution in Ontario (MNR 2009). While the forest-tundra ecotype also occurs in the province, the Range Management Approach focuses on the forest-dwelling ecotype, due to its threatened designation under Ontario’s Endangered Species Act, 2007. In order to clarify the distinction between forest-dwelling and forest- tundra woodland caribou and assess current approaches for delineating caribou ranges in the Far North of Ontario, researchers focused on characterizing differences in the behaviour and geographic distribution of the two ecotypes.
The CCP identifies preliminary ranges within the Continuous Distribution. Several different research efforts have been undertaken to determine whether or not there is evidence for the existence of distinct local caribou population ranges across the Continuous Distribution (Appendix 3-1). The research results can be used to:
- Assess the appropriateness of current approaches to caribou range delineation and
- Determine the degree of spatial discreteness and genetic connectivity that exists between local
- Identify landscape attributes that might influence the degree of movement and genetic connectivity between caribou
- Explore the underlying processes that drive caribou population distribution and population
- Determine whether landscape-level environmental variation creates population structure and geographic variation in potential adaptive behaviour
Methods & findings
Can the two caribou ecotypes be distinguished based on behavioural traits?
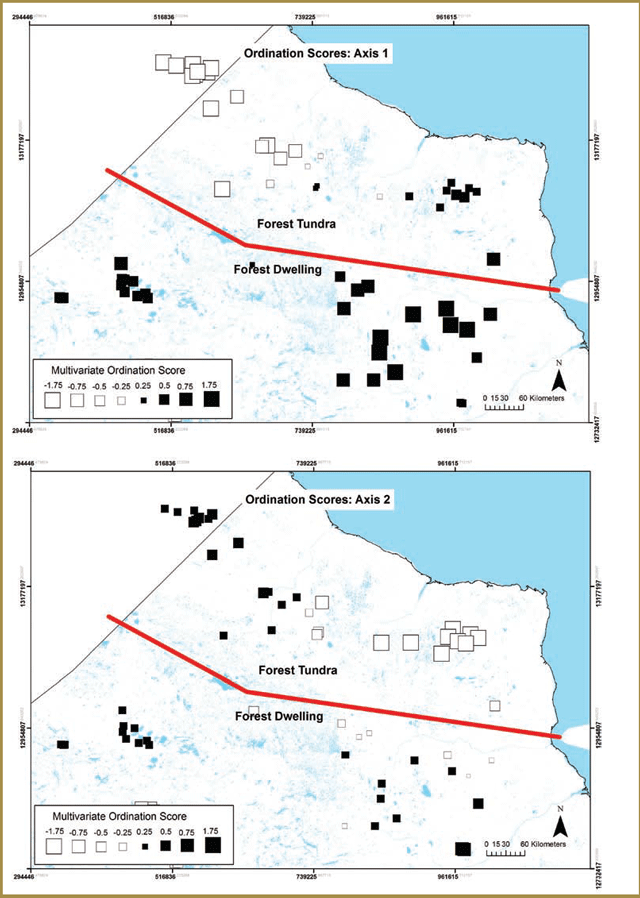
Two groups of researchers focused on analyzing geographic differences in individual movement and space use metrics to determine whether a clear behavioural and geographic distinction could be made between the two woodland caribou ecotypes [Project # 1, 2. Please see Appendix 3-1]. Both research groups analyzed caribou telemetry data collected as part of recent research as part of the monitoring and assessment activities (see Part 2 for more details) including the Far North Caribou Project and Integrated Range Assessments. Results from both projects [Project # 1, 2. Please see Appendix 3-1] provide support for the hypothesis that there is a geographic and behavioural distinction between the forest- dwelling and forest-tundra woodland caribou ecotypes.
One group looked at geographic differences in woodland caribou mobility and investigated whether there is a latitude- based threshold in general movement behaviour that might help to quantitatively demarcate the location of a transitional zone between the two ecotypes [Project # 1. Please see Appendix 3-1]. Their analysis of monthly movement rates showed that caribou at lower latitudes are less mobile and identified a distinct transitional breakpoint from relatively low to high movement rates at a latitude of approximately 53.7°N (decimal degrees) (Figure 3-2).
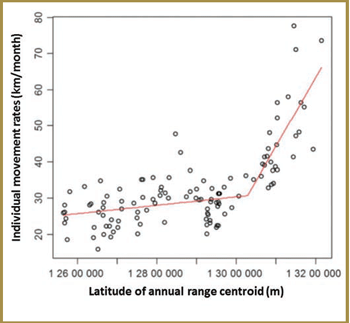
Figure 3-2. Latitudinal shift in caribou movement rates in Ontario: the transition from the forest-dwelling to forest-tundra ecotype (Figure 3 in Avgar et al. 2013). The X-axis values correspond to the projection used in the original analysis (i.e. Lambert Conformal Conic - LCC) – the breakpoint between the two ecotypes occurs at 13 028 647m (LCC) – this corresponds to a latitude of approximately 53.7°N (decimal degrees).
If you need alternate formats or communication supports to access this, please contact us.
Another group of researchers sought to identify the key behavioural indicators that best discriminated between the two ecotypes by performing metric-by-metric evaluations of caribou movement behaviour and geographic location [Project # 2. Please see Appendix 3-1]. Movement metrics included path lengths and the size and shapes of areas used by individual woodland caribou. Location attributes included minimum distances to key ecoregions and amount of time spent in the Hudson Bay Lowlands Ecozone. For each woodland caribou included in the analysis, metrics were measured at annual and seasonal (e.g. each calving season) time scales.
Analyses of spatial variation in these metrics detected a clear behavioural separation between the two ecotypes, but results also suggested that for part of the year (e.g. winter) there is a geographic overlap in the areas used by both ecotypes. A reduced number of key indicator variables that best distinguished between the two woodland caribou ecotypes were identified. While forest-dwelling caribou generally had smaller movement paths and annual and seasonal home range sizes than forest-tundra caribou, the three metrics that most clearly discriminated between the two ecotypes were all measured during the calving season: calving area perimeter, minimum distance to the Hudson Bay coast at calving and number of locations recorded in the Hudson Bay Lowlands during calving (e.g. Figure 3-3).
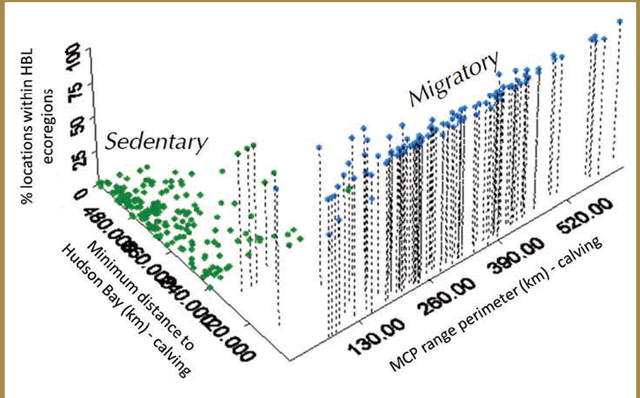
Figure 3-3. Ecotype indicator values for forest and forest- tundra woodland caribou in the Far North of Ontario (adapted from Figure 22 in Berglund et al. 2014). Each point indicates the corresponding values for the percentage of locations with the Hudson Bay Lowlands Ecozone, vs. minimum distance to the Hudson Bay coast during calving vs. calving area perimeter for a single woodland caribou during a single calving season. Green points represent the observations of caribou classified as forest-dwelling and blue points represent the observations of caribou classified as forest-tundra.
If you need alternate formats or communication supports to access this, please contact us.
The relative strength of these indicators reflects the higher degree of behavioural and geographic separation that exists for the two ecotypes during calving. During other times of the year (e.g. winter), there can be substantial spatial overlap in the areas used by forest-dwelling and forest-tundra woodland caribou (see Figure 3-6 in Section 3.2.2).
Is there evidence for local caribou populations?
In addition to evaluating evidence for the ecotype distinction, researchers have also examined whether there is evidence for local population structure amongst woodland caribou in Ontario [Project # 3, 4. Please see Appendix 3-1]. One research group analyzed a limited and patchy historic caribou telemetry dataset for caribou [Project # 3. Please see Appendix 3-1], which was collected at the southern margins of the Continuous Distribution from 1995 to 2008, prior to the initiation of data collection efforts related to Integrated Range Assessments. They focused on determining whether there was spatial clustering in the seasonal locations of caribou and assessing the degree of spatial separation between areas used by adjacent clusters of caribou.
Results indicated that even when patchiness of data collection efforts was accounted for, there was still some evidence for geographic groupings amongst caribou in areas along the southern boundary of the Continuous Distribution (Figure 3-4). However, the close proximity of boundaries of areas used by adjacent groups of animals indicates that while somewhat distinct, these groups are not discrete or isolated from each other (Figure 3-4). Therefore, caribou clusters in the southern portion of the Continuous Distribution are most appropriately characterized as demographic units (i.e. groups of individuals that are more likely to interact with each other and to be subject to shared resources, conditions and threatening processes) as opposed to discrete populations.
Another researcher analyzed the more comprehensive telemetry dataset collected across Ontario as part of recent monitoring and assessment and research efforts, in order to determine whether there are geographic differences in caribou behaviour and whether these differences correspond with spatial variation in different environmental characteristics [Project # 4. Please see Appendix 3-1]. Several behaviours and geographic characteristics were analyzed to identify spatial groups of caribou and determine whether there was evidence for population structure amongst and between forest-dwelling and forest-tundra woodland caribou. These included habitat selection during calving (e.g. use of shorelines, islands, and different forest types), seasonal migration distance, movement rates and fidelity to calving/rutting areas.
Two separate analyses were conducted – one for caribou distributed across the Continuous Distribution and one for forest-dwelling and forest-tundra woodland caribou in the Far North of Ontario. Preliminary results identified at least five spatial groups of caribou at the southern margins of the Continuous Distribution and at least three to four additional groups in the Far North of Ontario (Figure 3-5). Proximity of calving sites to lakes/islands, degree of selection for sparse conifer forest and treed wetlands, and movement rates are among the characteristics that make the greatest contribution to group distinctions. In areas where telemetry data coverage overlaps, the geographic divisions between caribou groups (Figure 8) correspond well with caribou clusters identified in the historic data analysis [Project # 3. Please see Appendix 3-1].
Preliminary results from an additional analysis of the behavioural characteristics of forest-dwelling and forest- tundra woodland caribou in the Far North of Ontario [Project # 4. Please see Appendix 3-1] indicated that while forest-dwelling caribou in this area are relatively similar, there is a distinction between the forest- dwelling and forest-tundra ecotypes (Figure 3-6). A possible East vs. West distinction between two groups of forest- tundra woodland caribou is also apparent. Preliminary results suggest that in comparison with forest-dwelling caribou, forest-tundra woodland caribou formed large aggregations near the coast of Hudson Bay during calving, exhibited lower selection for sparse conifer forest during calving, had greater directional persistence in movement paths and greater migration distances between summer and winter ranges.
Finally, caribou blood and fecal pellet samples have been analyzed to determine whether there are genetic differences amongst caribou distributed across Ontario (i.e. within and between ecotypes) [Project # 5. Please see Appendix 3-1]. Results obtained from genetic analyses are outlined in Part 2, Section 2.7.
Interpretation of research results
An improved understanding of the distinction between the two woodland caribou ecotypes that occur in Ontario helps clarify their spatial relationship and illuminate the level of interaction and behavioural differences that exist between them. It also informs efforts to delineate a geographic boundary between the two ecotypes. Likewise, information on current population structure is critical for delineating appropriate ranges for caribou as described in the Delineation of Woodland Caribou Ranges in Ontario (‘Range Delineation Report’) (MNRF 2014a). Research in both of these areas supports the implementation of the Range Management Approach prescribed in the CCP (MNR 2009 Action 2.0).
In addition to informing the range delineation process, evidence for substantial overlap in the northern-most areas used by forest-dwelling caribou and the southern-most areas used by forest-tundra caribou indicates that at certain times of the year (i.e. winter), it is not possible to make a geographic distinction between members of the two ecotypes. The lack of obvious morphological differences between the ecotypes contributes to this diffi . The existence of ecotype overlap should be considered when interpreting winter survey results in the overlap area. Evidence for the distinction between the forest-dwelling and forest-tundra ecotypes [Project # 1, 2, 5. Please see Appendix 3-1] and spatial structuring in caribou behaviour [Project # 4. Please see Appendix 3-1] was considered and incorporated in the range delineation process for ranges in the Far North of Ontario.
The delineation of the preliminary ranges in the CCP occurred prior to the completion of the research described here. However, Research results suggest that there is a good correspondence between the current seven more southern ranges (as described in the Range Delineation Report) and groups identified by analyzing historic [Project # 3. Please see Appendix 3-1] and recently collected telemetry data [Project # 4. Please see Appendix 3-1]. Specifically, there was fairly strong evidence for demographically and behaviourally distinct groups in the Wabakimi Provincial Park, Geraldton, Hearst and Cochrane areas. The geographic distinctions between the different groups are broadly comparable to the range boundaries [i.e. Brightsand (eastern boundary), Nipigon, Pagwachuan and Kesagami (i.e. western boundary with Pagwachuan)].
In some cases, the telemetry-based research results do not correspond as closely with the current delineation of ranges. These include:
- The lack of a clear distinction between caribou inhabiting the Brightsand, Churchill Ranges and Kinloch
- Evidence for behavioural and environmental distinctions between Lake Nipigon caribou and those inhabiting the surrounding
- Evidence for behavioural and environmental distinctions amongst a smaller sub-grouping of animals currently assigned to the Kesagami
Several of these patterns are further supported by results of genetic research [Project # 5. Please see Appendix 3-1] (see Part 2, Section 2.7). The interconnectedness of the central portion of the Continuous Distribution should be considered when conducting assessments of population and determining range condition. Maintaining the connectivity of suitable habitat across range boundaries supports the similarities amongst animals in the central portion of northwestern Ontario.
3.2.3 Identifying best population and health measures
Research indicates that direct estimation of caribou population size may be feasible in some situations; however, alternative approaches (like estimation of survival rates) may be more effective in other situations.
Future efforts to estimate caribou population size or map areas where they are likely to be found would benefit from selection of an appropriate sampling or survey design.
Research suggests that the quality of information obtained during aerial surveys could be improved by measuring variables that could affect observers’ abilities to detect caribou during surveys (like weather conditions) and using analysis techniques that directly account for the influence that survey conditions and the spatial interdependence of caribou observations can have on results
Research goals & objectives
As part of the CCP, Ontario committed to expand caribou monitoring efforts and develop standard protocols for collecting data on metrics like population size, range occupancy and population health (MNR 2009, Action 1.4). Two research projects have evaluated and developed alternative approaches for assessing population state and caribou occupancy patterns. One study evaluated the accuracy and cost-effectiveness of different methods for estimating caribou population size. The other focused on identifying landscape attributes that influence the occupancy patterns of caribou, moose and wolves, and applying a new analytical technique to improve the accuracy of occupancy models.
In addition, as part of the Research Program, several different measures of caribou population state and animal health were analyzed. Population state metrics included rate of population change (λ), as well as pregnancy, survival and recruitment rates. Caribou health was also measured at the individual level and included both long and short term indices of body condition. While research has focused on how these metrics are influenced by forest conditions and different levels of disturbance, researchers also gained insights into the relative advantages and disadvantages of using these metrics as measures of population status and health.
Methods & findings
What are the best approaches for estimating population size?
Researchers evaluated the costs and quality of results obtained using three different population estimation techniques, which were applied to a relatively isolated, high density caribou population (i.e. the Slate Islands caribou population) [Project # 6. Please see Appendix 3-1]. The three methods tested were winter aerial surveys using Forward Looking Infrared (FLIR) equipment, summer ground-based surveys and genetic analysis of fecal pellets collected during winter. Each technique produced different population size estimates for the Slate Islands caribou population. However, the specific estimates associated with each method fell within the relatively large confidence intervals associated with the population size estimates produced by all methods. The FLIR survey and the genetic sampling technique (with three sampling periods) produced the most precise population estimates. However, the genetic sampling technique can be more challenging when survey areas are larger and populations are not closed to immigration or emigration of new caribou.
Advantages associated with the FLIR approach included higher caribou detection rates than standard aerial census approaches and less observer bias (e.g. due to experience, fatigue, air sickness), while disadvantages include the effects that topography and dense conifer cover can have on animal detectability and the possibility of undercounting or double- counting animals.
What are the best approaches for modeling caribou occupancy in the far north of Ontario?
In a second study, researchers applied new analytical techniques to caribou, moose and wolf observations collected during systematic aerial surveys that were conducted in the Far North of Ontario and used them to develop occupancy models for each species [Project # 7. Please see Appendix 3-1]. The factors with the greatest impacts on animal detection, varied between species and ecozones (i.e. the Ontario Shield and the Hudson Bay Lowlands). In both ecozones, caribou were more likely to be detected when terrain openness was high. In the Hudson Bay Lowlands Ecozone, caribou detection was also influenced by time of year and time of day. Detection probability was highest earlier in the winter and at mid-day (vs. early or late in the day). Additionally, using an analytical technique that explicitly accounted for the lack of spatial independence between sampling locations improved the accuracy of occupancy models and the uncertainty associated with occupancy estimates.
What are the best approaches for measuring population trends and health?
The various population and individual condition metrics measured as part of the Research Program were derived in several different ways. Efforts to collect and analyze these metrics to test competing hypotheses about the factors affecting caribou persistence also highlighted their respective value and shortcomings for population status and health assessment [Project # 8. Please see Appendix 3-1]. These methods, along with the advantages and disadvantages associated with each measure, are outlined in Appendix 3-3. The general findings (i.e. study area comparisons) associated with the population status and health metrics measured as part of the Research Program, will be described in detail in upcoming sections.
Interpretation of research results
Population size can provide important insights into population status and viability, particularly when it is measured consistently over long periods of time. However, there are a number of difficulties associated with applying standard population estimation methods to wide-ranging, low density, difficult-to-detect species like caribou. The results of the population estimation study [Project # 6. Please see Appendix 3-1] (described above) highlight the costs and benefits of different techniques and illustrate the potential value of some innovative methods that could serve as improvements over traditional estimation approaches. The final selection of survey methods should be based on the specific monitoring/research questions being asked and the available resources. Ground-based surveys may be appropriate if a rough estimate of population size is sufficient, but it may be necessary to undertake more expensive surveys if more accurate and/or precise estimates are needed. In these cases, the FLIR method or the genetic sampling approach (using three sampling periods) should be considered. The use of multiple techniques can help mitigate the costs associated with the use of one technique alone and help improve certainty around estimation of population size.
Results from the occupancy modelling can be used to map the distribution of caribou across the landscape and the probability of caribou occurrence at different locations. As such, it can serve as a source of information for land use planning, the development of conservation strategies and long-term monitoring efforts (see Part 2, Section 2.5). Results from caribou occupancy modelling efforts in the Far North of Ontario [Project # 7. Please see Appendix 3-1] also demonstrate that it is important to account for the effects of factors that can influence the probability of detecting caribou during occupancy surveys and the lack of spatial independence in survey-based observations. The results also provide insights into specific factors or detection covariates that can affect the ability of observers to detect caribou (i.e. terrain openness, time of year and time of day) and demonstrate that their influence can vary between species and regions.
Future population estimation and occupancy surveys would benefit from efforts to account for factors that might hamper caribou detection by choosing an appropriate survey design (e.g. choice of survey timing, use of FLIR methods), measuring detection covariates and using analytical techniques that account for the influence of detection covariates and spatial dependence in survey observations.
Collection of population status and condition data as part of the Research Program [Project # 8. Please see Appendix 3-1] provided insight into caribou population status, adult and calf health and the different factors that might be influencing them. However, short- term measures of population state and animal health can be subject to considerable inter-annual variability, which can make it difficult to determine whether they are representative of long-term trends. Additionally, survival or recruitment rate estimates that are based on small samples of individuals are associated with a considerable degree of uncertainty, which can make it difficult to formulate a clear assessment of population state. Thus, while limited resources might favour less expensive methods and shorter and/or less extensive sampling regimes, it is important to recognize that choosing these options will result in trade-offs with respect to the quality and usefulness of results.
For example, calculations of intrinsic rate of population change (l) are based on multiple vital rate estimates (i.e. adult survival rates and recruitment rates), each of which is subject to environmental variation and is associated with its own level of uncertainty. Consequently, recommendations in favour of longer-term studies with large sample sizes are particularly important for this metric, which is difficult to derive with accuracy and precision. These difficulties suggest that l may be less valuable as an absolute measure of population status (especially when measured only once or for only a few consecutive years) and more useful as a relative metric that can be used to compare the impacts that alternative management scenarios might have on the long- term viability of a particular caribou population. Section 3.2.13 provides more details on this type of approach. In general, monitoring and assessment efforts might benefit from choosing a set of population and health metrics that are;
- Effective for measuring all characteristics of interest.
- Complementary, in that they compensate for the disadvantages associated with other metrics in the set.
3.2.4 Characterizing caribou habitat
Research results suggest that the habitat caribou selected and the amount of time caribou spent in different areas had a lot to do with food availability and avoiding areas where they were more likely to encounter predators.
Results from the research indicated that coniferous forests (particularly black spruce-dominated areas) and treed lowlands had year-round importance for caribou.
Research results indicated that caribou generally avoided disturbed habitats, areas with lots of deciduous trees, settlements, roads and open areas, but the extent to which they avoided these areas varied according to the season.
Research indicates that individual caribou returned to the same general areas they used in previous years, and this is especially true for areas where they gave birth and raised their young. Individual caribou were less likely to return to winter areas used in previous years, but their tendencies to return increased when they lived closer to roads and recently disturbed forests.
Research goals & objectives
Habitat can generally be defined as the combined set of resources and environmental conditions that result in the presence, survival and reproduction of a given organism (Caughley and Sinclair 1994). Ontario is committed to retaining caribou habitat in the amount and arrangement needed to sustain viable caribou populations within ranges (MNRF 2014b). To this end, the Boreal Landscape Guide (OMNR 2014) provides specific direction for identifying and managing caribou habitat at the landscape, stand and site scales during the forest management planning process. This direction was developed based on the best scientific information available at the time of guide development. In order to evaluate support for this direction and identify potential areas of improvement, there are several different caribou habitat research efforts that have been completed or are currently underway in Ontario (see also Section 3.2.14). All include direct assessments of the relationship between caribou use and the surrounding landscape and all are driven by the same basic goal - to improve our understanding of what constitutes caribou habitat at multiple scales.
Methods & findings
Which habitats do caribou use when feeding in managed and unmanaged forests?
Video data from GPS-collared caribou was analyzed to identify characteristics associated with communities caribou feed in. This method recorded evidence of the associated forest type and forage availability [Project # 9. Please see Appendix 3-1]. Preliminary results indicated that selection for feeding site habitats varied somewhat (as suggested by the variety of plants selected – see Section 3.3.4), but the majority of time spent feeding was in upland and lowland sites dominated by black spruce forest cover. During the summer, considerable feeding also occurred in herb-rich black spruce mixedwood sites.
What forest types do caribou use during summer?
To identify what types of forest stands are selected by caribou during the calving and post-calving seasons, researchers used location data from GPS-collared caribou to identify stands used by caribou during summer and compared used stand types to available stands across the broader landscape [Project # 10. Please see Appendix 3-1]. Preliminary results indicated that caribou in managed and unmanaged landscapes select conifer-dominated and treed lowland stands, while avoiding disturbed areas and deciduous stands. Unlike caribou living in the unmanaged landscape, caribou in the managed landscape selected sparsely treed stands and avoided open lowlands. They also exhibited stronger selection for conifer but weaker avoidance of deciduous stands.
How is caribou movement behaviour affected by different landscape attributes and environmental conditions?
To determine how caribou movement rates change in response to different resources and conditions, researchers used location data from GPS-collared caribou to analyze how caribou movement patterns and step-based selection of habitats (which compares actual to potential movement steps) are influenced by variation in food availability, snow depth, roads and the probability of habitat use by wolves and moose, all of which are influenced by forest stand characteristics shaped by natural and human disturbance [Project # 11. Please see Appendix 3-1]. Researchers also developed a novel approach for modeling caribou movement, in which simulated individuals have sensory, memory and movement capacities and different strengths of attraction or repulsion to different landscape attributes in different seasons [Project # 49. Please see Appendix 3-1] (see Section 3.2.13).
Analyses of caribou movement patterns indicate that a great deal of observed variation in caribou movement behaviour can be attributed to local landscape structure and environmental conditions. Monthly measures of distance traveled and movement path linearity suggest that caribou respond strongly to local variation in both predation risk and forage availability, by choosing to spend most of their time in areas with higher vegetation cover (in summer and winter) and conifer forest (in winter). Most analyses indicate caribou avoidance of early successional stands, which are commonly used by moose (see Section 3.2.9), as well as areas with extensive linear features (e.g. roads) that are heavily used by wolves (see Section 3.2.10). The only exception to this was some evidence for relatively weak selection of regenerating and deciduous stands in winter. Dominant patterns of caribou habitat selection likely serve to reduce exposure to predation risk to some degree.
Caribou selected forest stand types that have the highest abundance of preferred diet items (i.e. ground lichens), which should serve to increase energy gain. During the summer, caribou spent more time in areas with considerable amounts of conifer cover and water, both of which are thought to provide refuge in this season. Preliminary results indicate that caribou whose home ranges have higher densities of wolves and moose tend to be more careful about choosing habitats to reduce that risk. In winter, caribou whose home ranges have lower food abundance tend to be more careful about selecting forest stands that offer the best foraging opportunities. Results from other analyses indicate that conditions related to ease of movement and exposure (i.e. open vs. closed cover) also affected caribou movement behavior [Project # 11. Please see Appendix 3-1] (Avgar et al. 2013). Caribou moved more when travelling through open habitats during summer and they moved less when snow depths increased during winter. Preliminary results also indicate that spatial memory increased the probability of re-use of particular forest stands, despite additional evidence that indicated that caribou perception of surrounding habitat conditions was limited to 100-200m [Project # 49. Please see Appendix 3-1].
Which broad-scale landscape attributes influence winter occupancy by caribou in the far north of Ontario?
To determine what general landscape attributes influenced the probability of caribou occupancy in the Far North of Ontario during winter, researchers examined the relationship between caribou observation data collected during systematic aerial surveys (see Section 3.2.3) and several different landscape attributes, including the amount of water, bogs and burned or harvested areas, terrain ruggedness and distance to settlements [Project # 7. Please see Appendix 3-1]. The probability of winter occupancy by caribou in both ecozones in the Far North of Ontario (i.e. the Hudson Bay Lowlands and the Ontario Shield) was lower when the amount of water was more extensive and became higher with increasing distance to nearest settlements. Other attributes associated with caribou occupancy differed between the two ecozones. In the Ontario Shield Ecozone, caribou occupancy increased as the amount of bog increased, but was lower when terrain ruggedness and disturbed habitat were more extensive. In the Hudson Bay Lowlands, caribou occupancy was lower in areas with more extensive bogs. Geographic variation in the distribution of the different landscape attributes that influenced caribou occupancy led to clear spatial patterns in occupancy across the Far North of Ontario.
Caribou occupancy was highest along the boundary between the two ecozones (i.e. the ecotone) and lowest in the north- east area of the Hudson Bay Lowlands, along the Hudson Bay coast and in the western Ontario Shield. A probability of occupancy map that depicts these results (in combination with probability of winter occupancy for Kinloch Range and the more southern ranges) is displayed in Figure 2-13 in Part 2, Section 2.5.
How faithful are caribou to seasonal home ranges and calving sites and is traditional use affected by different landscape attributes?
To determine whether landscape attributes affected the tendency of individuals to return to the same seasonal home ranges [Project # 12. Please see Appendix 3-1] and calving sites [Project # 13. Please see Appendix 3-1] used over multiple years, researchers used telemetry data from caribou that were GPS- collared for more than one year, along with spatial data on forest type and natural and human disturbance. Preliminary results from analyses of seasonal home ranges indicated that traditional use (i.e. fidelity) to areas used within seasons was lowest during winter and during the month prior to calving [Project # 12. Please see Appendix 3-1]. However, it increased at calving and was even higher during the post-calving season. Landscape characteristics had little influence on fidelity by individual caribou in all seasons except winter, where traditional use of seasonal home ranges increased when caribou were closer to roads and young forests. In contrast, fidelity to areas used during winter decreased when caribou were closer to mature upland forests.
To investigate finer scale traditional use of key sites (i.e. calving sites), researchers used marked and prolonged declines in caribou movement rates to identify calving events and determine the geographic location of forest-dwelling and forest-tundra woodland caribou at the start of calving [Project # 13. Please see Appendix 3-1]. Once calving sites were identified, they measured the distance between calving sites used by individual caribou in multiple years. Results indicated that the distance between calving sites in successive years was consistently smaller for the forest-dwelling ecotype (median distance: 10km) than it was for the forest-tundra ecotype (median distance: 50km), which suggests that there is some degree of calving site fidelity for the forest-dwelling ecotype. Forest-tundra woodland caribou did not exhibit the same level of absolute site fidelity, but given the much larger sizes of both their home range and distances they travel to reach their calving grounds, distances between successive calving sites might also be interpreted as indicating relatively high fidelity to these sites.
Are caribou home range sizes affected by different landscape attributes and environmental conditions?
To determine whether individual home range sizes were influenced by landscape attributes and environmental conditions, researchers generated individual home ranges using caribou telemetry data and analyzed whether home range sizes were influenced by a variety of different factors (e.g. forest or lowland community type, density of edges/ transitional zones between communities, weather) [Project # 14. Please see Appendix 3-1]. Results indicated that the areas used over the course of a year by both forest-tundra and forest-dwelling caribou were larger when the amounts of conifer forest and treed wetland cover were low. Larger home ranges were also associated with high amounts of wetland edges, exposed ground and mosses and lichens.
Interpretation of results
The new insights obtained from research into different aspects of caribou habitat selection can contribute to efforts to identify suitable caribou habitat. Greater understanding of what motivates caribou movement and habitat selection at different scales (i.e. forage acquisition vs. predator avoidance) can also be useful for determining how caribou are likely to respond to and be affected by disturbance. Each of the different habitat-related research projects has different implications for understanding caribou ecology (also see Appendix 3-4 for a list of relevant projects currently underway).
While there was some variation in the forest types that caribou selected for feeding in different seasons, preliminary results illustrated the year-long importance of coniferous forests (particularly black spruce-dominated stands) as a food source [Project # 9. Please see Appendix 3-1]. Analyses of caribou movement and space use behaviour at multiple scales and different seasons [Project # 7, 10, 11. Please see Appendix 3-1] also provided support for selection of conifer forests and treed wetlands by caribou and avoidance of disturbed habitats, settled and open areas and roads (see Section 3.2.14 for additional evidence for these selection patterns). Preferred forest types provide higher than average food and energetic availability (see Section 3.2.7), as well as lower risk of predation [Project # 11. Please see Appendix 3-1] (also see Section 3.2.9).
Research into how caribou movement patterns respond to different resources and environmental conditions [Project # 11. Please see Appendix 3-1] has provided some unique insights into what factors influence caribou habitat selection during summer and winter seasons. Demonstrating that in addition to habitat characteristics (e.g. forest type, roads) that are more fixed in space and time, local environmental conditions such as predation risk, exposure to apparent competitors like moose and food availability (which can be more dynamic in space and time) can have a strong effect on how long caribou spend in an area. The approach for modeling caribou movement responses to different resources and conditions, developed as part of this research, is also serving as a framework for representing caribou movement in a spatial Population Viability Analysis (PVA) that has been developed for caribou (see Section 3.2.13).
Research on the influence of different resources and conditions on home range size [Project # 13. Please see Appendix 3-1] and caribou fidelity to seasonal areas [Project # 12. Please see Appendix 3-1] has implications for the effects habitat quality might have on caribou condition and population viability. When cover types with high habitat value are abundant (e.g. conifer forests and treed lowlands), caribou travel within smaller annual areas (i.e. home ranges) than they do when the amount of good habitat is low. This suggests that caribou might be expending more energy to meet their needs when the abundance of suitable habitat is low and they may also have a higher probability of entering risky habitats while travelling through a large home range. This could have a negative effect on their energy balance (see Section 3.2.4), body condition, survival and reproductive success.
Finally, research on the traditional use seasonal ranges and calving sites indicates that individual caribou exhibit a varying degree of between-year fidelity to areas used at different times of the year. Forest-dwelling caribou in particular, exhibit a high degree of fidelity to calving and post calving areas [Project # 12. Please see Appendix 3-1], as well as specific calving sites [Project # 13. Please see Appendix 3-1]. However, preliminary results from analyses conducted at the seasonal range scale, indicated that with the exception of winter ranges, associated forest and disturbance types had little influence on multi-year use of traditional seasonal ranges by caribou [Project # 12. Please see Appendix 3-1]. The tendency of individuals to maintain fidelity to familiar seasonal sites, regardless of associated habitat suitability, may be a maladaptive trait (Faille et al. 2010). In other words, tendencies to return to familiar sites could lead caribou to use areas with characteristics that could be detrimental to their probability of survival and/or reproductive success (e.g. areas with high densities of roads, recent cuts) (Faille et al. 2010). Preliminary evidence for increased winter range fidelity amongst caribou that are close to roads and recently disturbed areas [Project # 12. Please see Appendix 3-1] suggests that in more disturbed landscapes, individual caribou may be more likely to restrict winter use to areas they are familiar with. If this leads to more intensive caribou use of specific winter areas at the population level, limited winter food availability might have a negative impact on caribou condition. Additionally, stronger traditional use of winter areas in landscapes with higher levels of disturbance [Project # 12. Please see Appendix 3-1] is also consistent with the caribou behaviour component of the Predator Escape Hypothesis (see Section 3.2.11). However, none of the evidence from other research projects supports the predator response component of this hypothesis – namely, that if caribou in managed landscapes are restricted to relatively small areas that receive consistent use, wolves will be better able to detect and target them (see Sections 3.2.9 and 3.2.10).
3.2.5 Enhancing regeneration of harvested areas
Research indicates that there were differences in the plant species that grew in previously harvested areas (in comparison with non-harvested areas). However, the amount, diversity and species composition of lichens growing in stands that were originally disturbed by harvesting, did not differ greatly from that of lichens growing in stands that were originally disturbed by fire.
Research suggests that when different types of silviculture treatments were used on previously harvested forests, they had different short and longer-term effects (both negative and positive) on lichen communities.
Researchers found that during summer, caribou tended to use areas with greater amounts of ground lichen, which is a forest characteristic that could be enhanced by applying different silviculture treatments.
Research goals & objectives
As part of a general commitment to enhance caribou science (MNR 2009), Ontario committed to undertake research on silvicultural efforts to promote the regeneration of caribou habitat following forest harvesting. There are several complete and ongoing research projects that focus on the regeneration of harvested areas. These research projects centre on two main goals. The first goal is to document the specific impacts of different disturbance types on the plant communities that caribou rely on for cover and food. Relevant research projects focus on determining how forest composition and structure changes in response to disturbance type and age. The second goal is to characterize the effects of stand-level silvicultural treatments on lichen – the main caribou food item (See Section 3.2.7).
Methods & findings
What effects do wildfire and harvesting have on the composition and diversity of vegetation communities?
To characterize the effects of different disturbance types (i.e. wildfire and harvesting), forest types and forest ages on plant diversity and composition, researchers sampled forested stands with different cover types, ages and disturbance histories [Project # 15. Please see Appendix 3-1]. Data on several attributes of vegetation communities, as well as soil and site conditions were collected and analyzed. Plant communities in natural origin stands differed from plant communities in harvested stands. The degree of difference between the composition of plant communities found in natural origin stands and the composition of plant communities found in harvested stands increased as forest age increased. In contrast, while plant diversity (i.e. the number and relative abundance of different plant species) was generally higher in harvested stands than it was in natural origin stands, the size of this difference decreased as forest age increased. Differences in forest structure between harvested and natural-origin stands were not as pronounced. Young managed and unmanaged stands differed in landform, organic layer depth and soil depth, but there were no differences between medium age forests. Amongst older forests, the key difference was canopy closure, which was significantly higher in harvested ( vs. natural origin) stands.
How do disturbance type, stand type and forest structure affect the abundance, biomass and diversity of ground lichens?
Researchers measured lichen abundance and a variety of different stand characteristics in natural origin and previously harvested forest stands to determine how disturbance type, forest type and forest structure affects the abundance and diversity of ground lichen [Project # 16. Please see Appendix 3-1]. The amount of lichen (abundance) was greatest in conifer-dominated stands ( vs. deciduous or mixedwood forests with non-organic soils). When forest and soil types were similar, there were no differences in lichen abundance between natural origin and harvested stands. Amongst stand types where lichens were most abundant (i.e. non-organic conifer-dominated stands), the amount of lichen increased with decreasing canopy closure, stand density, tree height and crown height.
To determine whether forest structure and age influence the diversity of ground and tree lichens, another group of researchers measured lichen composition, lichen biomass and different attributes of forest structure in harvested and natural-origin conifer forests [Project # 17. Please see Appendix 3-1]. Results suggest that the number of lichen species grew in association with increases in the number of fine-scale microhabitats within a stand, variation in canopy closure (and light conditions) and stand age. While the structural complexity of forests often increases with age, older stands with closed canopies were less likely to have a large number of lichen species due to insufficient light. Lichen biomass increased with increases in light penetration, stand age and the sandiness of the soil.
What are the impacts of different silvicultural treatments on ground and tree lichen communities?
Researchers compared the abundance of ground lichens in 20 to 40 year-old stands that were regenerating after the application of one of two silvicultural treatments: prescribed burning or mechanical site preparation [Project # 18. Please see Appendix 3-1]. Results indicate that while lichen abundance was generally low in stands exposed to each treatment type, there was a trend towards higher abundances in stands treated with prescribed burns vs. those treated with mechanical site preparation. Additional data has been collected by MNRF and industry partners in stands outside the original study areas, and will be incorporated into further analysis of this question.
To determine whether different herbicide treatments had short-term impacts on the diversity of ground and tree lichens, researchers applied high and low concentrations of different herbicides (Triclopyr and glyphosate) in plots established throughout a black spruce and jack pine- dominated forest with extensive lichen cover [Project # 19. Please see Appendix 3-1]. The abundance and composition of different lichen species was measured and compared before herbicide application and one year after. Lichen abundance was reduced by 40% (for Triclopyr) and 56% (for glyphosate), with the greatest impacts observed for lichen species with heavily branched structures.
Researchers also studied the long-term impacts of herbicide applications and other silvicultural treatments on the diversity of ground and tree lichens, by measuring lichen composition and biomass in harvested and natural-origin conifer stands that were 25 to 40 years of age [Project # 20. Please see Appendix 3-1]. Harvested stands had been treated with 1 of 3 treatments: harvested and planted; harvested, planted and treated with glyphosate herbicide; harvested, planted and treated with a different herbicide (2, 4-D). Stands treated with herbicides had lower lichen biomass and different community structure than both harvested and planted stands (with no herbicide application) and natural origin stands. The composition of lichen communities varied amongst natural origin and harvested and treated stands, but there was considerable overlap, except for stands treated with 2,4-D, which contained lichen communities that were notably different from those observed in other stands.
Are there forest characteristics associated with caribou summer use that could be influenced by silvicultural techniques?
Researchers used location data from GPS-collared caribou to identify stands used by caribou during summer [Project # 10. Please see Appendix 3-1]. Numerous used sites in managed and unmanaged landscapes were sampled to collect vegetation and stand structure data (e.g. tree density, visibility). Analyses focused on identifying stand structural characteristics that are associated with caribou use in preferred stand types (i.e. conifer dominated stands and treed lowlands – see Section 3.2.4 [Project # 10. Please see Appendix 3-1]). Preliminary results indicate that when conifer-dominated and treed lowlands stands used by caribou were compared to available stands, most of the stand characteristics did not differ. However, there were a few notable exceptions [Project # 10. Please see Appendix 3-1]. Used conifer and treed lowland stands in both managed and unmanaged landscapes contained more ground lichen biomass than available stands with the same cover type. Additionally, in the managed landscape, canopy closure was lower in used conifer stands than it was in available conifer stands, but no such difference was documented in the unmanaged landscape.
Interpretation of research results
In recent years, emulation of natural disturbance patterns has been one of the primary goals of forest management planning and practice (MNR 2001; OMNR 2014). It is believed that efforts to minimize differences between natural origin and harvested landscapes will maintain adequate food and cover for caribou and other boreal species.
Research results suggest that there are significant differences between vegetation communities in harvested and natural origin stands. Differences most relevant to caribou conservation are likely those that affect the abundance, biomass and diversity of lichen, which is the main year-round food source for caribou (see Section 3.2.4). The results of one study [Project # 15. Please see Appendix 3-1] suggested that older harvested stands have higher canopy closure than older natural origin stands, which could result in less light penetration and lower lichen abundance. However, results from direct research into differences between lichen communities in managed vs. unmanaged stands, documented no significant differences when stand type, age and soil conditions were similar [Project # 16. Please see Appendix 3-1]. Yet, all lichen-focused studies described here suggest that conifer stands have higher abundances of lichen than other stand types. Where harvesting produces landscapes with less conifer forest and more mixedwood and deciduous cover, the amount of lichen biomass at the landscape scale could be lower than what occurs in unmanaged landscapes.
In addition to coniferous tree cover, other stand attributes positively associated with lichen abundance and diversity include non-organic sandy soils, along with several characteristics often associated with greater stand age (i.e. lower tree density, greater canopy openness and greater tree and crown height). Results suggest that if conditions are appropriate (e.g. for sites with non-organic, sandy soils), silvicultural treatments aimed at reducing tree density and increasing canopy openness may produce higher lichen abundance and better emulate the stand structure found in natural origin forests.
Several of the studies described in this Section also evaluated the short and long-term effects of different silvicultural treatments on lichen communities. The results provide insight into the effectiveness of different treatments for emulating natural disturbance and promoting lichen growth. For example, when compared with mechanical site preparation, prescribed burning is associated with higher ground lichen abundance [Project # 16. Please see Appendix 3-1]. Drawing clear conclusions about the impact of short and long-term effects of herbicides on lichen is complicated.
Without applying silvicultural treatments, stands that re- grow after harvesting are less likely to be conifer-dominated than they would have been after a natural wildfire. As a result, tree-planting, herbicide applications and other silvicultural measures are often applied to control deciduous re-growth and prevent harvested stands from converting to mixedwood. While herbicides help regenerate stands with characteristics that are important for lichen growth (i.e. conifer-dominated tree cover), they also appear to have negative short and longer-term impacts on lichen abundance (and in some cases, composition) when compared to harvested stands that were planted but had no herbicide applications and natural origin stands. These results suggest that lower impact herbicides (e.g. Triclopyr vs. glyphosate or 2, 4-D) and/or reduced applications may help regenerate caribou habitat (McMullin et al. 2013). Leaving untreated patches in larger sprayed areas is one herbicide reduction technique that might facilitate re-colonization by lichens.
Finally, preliminary results from the evaluation of stand structural characteristics associated with caribou use have provided insight into characteristics associated with caribou occupancy of selected stand types during the calving and post-calving seasons (i.e. higher lichen biomass in conifer and treed lowland stands and lower canopy closure in conifer stands) [Project # 10. Please see Appendix 3-1]. Identifying these characteristics provides insight into the motivations that might drive caribou occupancy of selected conifer or treed lowland stand types (e.g. food availability). Additionally, since these characteristics can also be influenced by different forest management techniques, these preliminary results provide insights into what stand characteristics are associated with caribou use during the calving and post-calving periods.
3.2.6 Caribou re-occupancy or use of formerly harvested habitats
Research indicates that caribou generally avoided areas that were recently harvested, but some infrequent examples where caribou used relatively young, harvest origin forest were documented. However, these occurrences are likely not good examples of caribou re-occupancy or use of habitat that is likely to improve their chances of surviving over the long-term.
Research goals & objectives
An improved understanding of how and when caribou re- occupy previously harvested areas can help inform efforts to regenerate suitable caribou habitat (MNR 2009 – Action 1.0). An objective of the Research Program was to identify examples of caribou re-occupancy or use (for cases when prior occupancy is uncertain) of harvest origin stands and determine what factors influence this re-occupancy or use.
Methods & findings
Researchers used location data from GPS-collared caribou to identify use of previously harvested stands during late spring and summer [Project # 21. Please see Appendix 3-1]. Characteristics of harvest origin stands that were either used by caribou or available on the landscape were sampled and compared in a single managed area to determine whether any attributes differed between them.
The number of previously harvested stands with evidence of caribou use was relatively small. A total of 16 previously harvested stands that were used by caribou were identified and sampled, then compared to 46 available stands that were also previously harvested. Preliminary results indicate that stands used by caribou varied from 8 to 68 years of age, but they tended to be younger (median age: 12 years) than other harvest origin stands (median age: 38 years). The stands used by caribou also tended to be conifer-dominated, with median conifer basal area (a measure of stand density) greater than 80%. Preliminary results also indicated that harvest origin stands used by caribou tended to contain shorter trees with smaller diameters than available harvest origin stands. They also had lower tree density and lower canopy closure. Research into caribou use of previously harvested stands is ongoing (see Appendix 3-4), although analyses have been hampered by the availability of accurate spatial data for harvested areas that includes information on date and method of harvest.
Interpretation of research results
Identifying re-occupancy of previously harvested areas is challenging (see detailed discussion in Section 3.3.4), but in general, results of research into caribou use of previously harvested areas can help identify characteristics associated with re-occupancy, provided those stands are old enough to have developed the general attributes associated with suitable caribou habitat. Better information about attributes that influence caribou use of previously harvested areas can improve harvesting practices. It can also be used to assess the effectiveness of different silvicultural practices for creating suitable caribou habitat.
The examples of caribou use of harvested stands that were identified in the research described here appear more representative of latent use of harvest origin stands in the early post-harvest period, rather than longer term re-occupancy of older, previously harvested stands that have regenerated into suitable habitat. Almost all of the harvest origin stands used by caribou were classified as disturbed using current provincial land cover data. The disturbed classification is intended to reflect relatively young regenerating stands (i.e. around 20 years or younger), but 4 of these 14 stands were older (> than 30 years).
Results from several other research projects that focus on caribou habitat selection have indicated that younger disturbed forests are either avoided or aren’t actively selected by caribou, particularly during summer (see Section 3.2.4). Other research results described here indicate that disturbed forests are also associated with higher levels of predation risk (see Section 3.2.9) and relatively low amounts of ground lichen (see Section 3.2.5). Research from other jurisdictions suggests that the use of such areas by female caribou may be maladaptive, particularly in summer (Dussault et al. 2012). Infrequent caribou use of young harvested stands likely is not representative of longer-term re-occupancy. In order to identify stand characteristics associated with caribou re-occupancy or use, more information is needed on use of older, suitable harvest origin stands (Appendix 3-4).
The preliminary results may not have value for understanding longer-term re-occupancy of harvested stands, but can provide some insight into potential motivations that might underlie late spring and summer use of young harvest origin stands when it occurs. For example, smaller tree diameters and lower tree density could provide greater ease of movement for caribou travelling through recently disturbed areas as they move between actively selected forest types. A further explanation for the relatively young age of used harvest origin stands includes feeding on sedges and grasses, which comprise a greater portion of caribou diets in managed landscapes (see Section 3.2.7) and are often abundant in recently harvested areas (e.g. Qi and Scarratt 1998, Martin-DeMoor et al. 2010 – cited in Thompson et al. submitted manuscript). The validity of both of these potential explanations remains untested.
3.2.7 The energetic balance hypothesis: Caribou diet, nutrition, energy expenditure and condition
Research indicates that ground lichens were the main sources of food for caribou throughout the year (including summer), although researchers also found that lichen is not very nutritious.
Research suggests that local environmental conditions such as food availability, snow depth and temperature affected how much energy caribou use. Distance travelled also affected caribou energy use.
Researchers found that biting flies (such as deer and horse flies) occurred in greater amounts in open harvested areas than they did in forested areas. Also, video analysis suggests that caribou may have spent less time feeding when there were a lot of insects harassing them.
Research into the body condition of adult caribou and calves suggests that the total amount of energy they get may have been lower when they lived in areas with considerable amounts of human disturbance.
Research goals & objectives
In an effort to understand the influence that energetic balance might have on caribou condition and vital rates, researchers involved in the Research Program have been trying to improve understandings of different aspects of caribou energy intake (through eating) and energy expenditure (e.g. through activities like running or walking) and determine whether these aspects are affected by human disturbance; and determine whether caribou condition and vital rates indicate that energetic balance might be affecting caribou health and population growth.
Methods & findings
What do caribou eat?
Caribou diet and how it varies throughout the year was characterized using two innovative methods; DNA analysis of fecal pellet contents and the analysis of very high resolution videos of caribou feeding activities [Project # 22. Please see Appendix 3-1]. Scat was collected from collared animals when they were captured in winter and video data were recorded in all seasons using cameras attached to specialized GPS-collars that were deployed on some collared caribou.
Results indicated that caribou winter diets are comprised of a very limited number of species: ground lichens (which comprise 65% of caribou winter diets), tree lichens and mosses. In spring and summer, the importance of ground lichens decreased and caribou diets broadened to include grasses and sedges, herbaceous plants, mosses and shrub leaves. However, ground lichens still made up over half of the total quantity of forage eaten by GPS-collared (video) caribou in early spring, late spring and summer, while only limited consumption of tree lichens was observed for any season.
There were some differences in caribou diets between managed and unmanaged landscapes. These differences were most apparent during summer when the diversity of forage species consumed was lower in the unmanaged landscape.
Grasses and sedges were more commonly eaten in managed landscapes during all seasons; however, most summer diet differences do not appear to reflect the higher level of anthropogenic disturbance in the managed study areas.
What is the nutritional value of different forage species?
To characterize the nutritional value of different forage species, researchers sampled forested sites containing a variety of different tree ages, disturbance histories, community types and substrates (i.e. upland vs. peatland) [Project # 23. Please see Appendix 3-1]. Data on plant diversity and different measures of nutritional value were collected at each sampled site. Results indicated that disturbance type (i.e. wildfire vs. forest harvesting) alone did not have a significant effect on plant biomass or productivity, but forest age and community type did. The biomass of species consumed by caribou responded differently to forest age and type. Lichen, grass and sedge biomass was lowest in older uplands (71+ years) and highest in younger stands (<30 years), while feather and sphagnum moss biomass was highest in older upland and lowland stands, respectively. Plant nutritional value was not affected by stand type or age, but it did differ substantially between species. Ground lichens and sphagnum mosses had the lowest nutritional value, while deciduous shrubs and herbaceous plants had the highest nutritional value. Overall, lowlands (vs. uplands) and younger stands (vs. intermediate age and older stands) contained plant communities with higher biomass, productivity and nutritional value.
What are the energetic costs of different activities and how are these affected by environmental conditions?
To measure the amount of energy expended by caribou during different behaviours and activities, researchers analyzed activity data recorded by accelerometers from GPS- collared caribou and European reindeer [Project # 24. Please see Appendix 3-1]. Researchers studied the relationship between different behaviours and activity levels amongst a small sample of captive European reindeer
Researchers also looked at how activity levels of caribou changed in response to changes in vegetation cover, temperature and snow depth. Results from captive European reindeer and field-based studies using GPS-collared (video) caribou documented a positive relationship between activity levels (measured using collar accelerometers) and caribou energy expenditures. Studies of collared caribou indicated that energy expenditures are heavily affected by individual movement rates, but they are also influenced to a lesser extent by a suite of local environmental conditions (including snow depth, amount of vegetation cover and temperature). Energy expenditures increased with higher movement distances, were highest at intermediate levels of vegetation abundance (likely due to foraging behaviour) and increased with greater snow depths and exposure to cold winter temperatures (likely due to increased locomotion and thermoregulation efforts).
Other researchers have undertaken a more detailed evaluation of the impact of snow conditions on caribou energetics by collecting and analyzing extensive field data on snow depth and structure in different forest communities [Project # 25. Please see Appendix 3-1]. Snow condition data were used to generate a model that predicts daily snow depth across the studied landscapes, incorporating differences between different forest types. Preliminary results from analysis of field data indicate that snow accumulation and melting are linked to canopy cover, with snow accumulating and melting more quickly in more open ( vs. high cover) habitats. Understanding and modeling general patterns of snow accumulation on lakes is more complicated due to slush formation and freezing processes and the shallower and denser snow cover that wind effects produce on lake surfaces.
How does forest age and disturbance origin affect biting insects and how do caribou respond to insect harassment?
Researchers have been undertaking different approaches to develop a better understanding of the impacts of insect harassment on caribou energetics. One group of researchers sampled biting flies (i.e. horse and deerflies, mosquitoes and black flies) from June to August in forests with different tree ages, disturbance histories and community types [Project # 26. Please see Appendix 3-1]. Preliminary results indicate that different fly families had different habitat preferences, but all three occurred in higher densities in young, recently harvested stands.
Another group of researchers analyzed videos from GPS- collared caribou (video) to document seasonal patterns in insect harassment and common caribou responses to the presence of different insect types and numbers [Project # 27. Please see Appendix 3-1]. Preliminary results from the analysis of video data from caribou collars indicate that insect harassment of caribou occurred from June-August (peaking in July) and when harassment levels were high, caribou appeared to spend less time feeding and moving and more time lying down. Additional preliminary results from the field study of biting flies [Project # 26. Please see Appendix 3-1] suggest that restricted movements by caribou could be a strategy to reduce exposure to new flies that haven’t fed yet.
Does caribou energetics differ between managed and unmanaged landscapes?
Researchers have been working to develop a comprehensive ecological energetics model for caribou [Project # 28. Please see Appendix 3-1] that integrates the results from other studies described in this Section. The general equation that has been developed to determine how much energy is available to caribou for growth and reproduction is:
Energy Available = Energy gained from food – (Basal metabolic rate + Energy used to feed + Energy used to move + Heat lost in wastes + Energy lost to avoid insects + Energy lost to regulate body temperature)
Two energetics models (for managed and unmanaged landscapes) are currently being constructed and compared. The values used to apply this equation are being derived from other Research Program studies.
Do caribou vital rates and body condition differ between managed and unmanaged landscapes?
Researchers have been using data obtained from collared caribou to determine whether body condition and some key vital rates known to be negatively influenced by poor nutritional status (e.g. pregnancy rates, live birth rates) differ between managed and unmanaged areas [Project # 29. Please see Appendix 3-1]. Data analyzed included body measurements and blood samples taken when collaring adult caribou and video evidence for calf birth rates, calf condition and timing of calf deaths. Calf condition was assessed using video evidence for body fat levels and behaviour.
Preliminary results from vital rate comparisons and body condition analyses provide some support for the predictions of the Energetic Balance Hypothesis. Pregnancy rates in all landscapes were relatively high (i.e. 0.80 to 0.89), but there was a trend towards higher rates in unmanaged vs. managed landscapes. In general, small sample sizes of GPS-collared (video) caribou prevent strong conclusions from being drawn about differences in calf birth rates, calf condition and timing of calf deaths between managed and unmanaged landscapes. However, some general patterns in were apparent. Preliminary results indicate that there are no apparent differences in live birth rates between study areas. Calf mortality rates, the timing of calf births (median date: May 21) and the timing of calf deaths (median dates: mid-late June) were quite similar across landscapes and between years. Most viable calves born in all study areas died before mid- July. Preliminary results from an analysis of calf condition, suggest that calves born in managed landscapes may be in poorer condition at birth than calves born in unmanaged landscapes but further analyses are needed to confirm this trend. Similar patterns were documented in analyses of adult condition. Preliminary results indicate that while adult condition in all study areas varied between years, adult caribou from managed landscapes were in poorer condition than those from unmanaged landscapes. Preliminary results also indicated that this managed vs. unmanaged difference was more pronounced for nutritional conditions experienced during winter than it was for summer nutritional conditions.
Interpretation of research results
Evidence that caribou diets broaden during summer to encompass a wider variety of plant types is not surprising. The fact that lichen remains the dominant year-round food item [Project # 22. Please see Appendix 3-1] was not expected and was previously unknown, except for a single study from Newfoundland (Bergerud 1972). Lichens are slow-growing, require certain soil conditions to grow, and can be eliminated or reduced by herbicides (see Section 3.2.5) and/or destroyed by equipment during summer harvesting. Thus, efforts to regenerate long- term caribou habitat in harvested areas should consider how to support lichen production.
The amount of general and plant-specific biomass available to caribou varies between community types (i.e. lowlands vs. uplands) and forest ages [Project # 23. Please see Appendix 3-1]. These results suggest that differences in landscape composition and/or habitat selected by caribou within a particular landscape, could affect the quantity and quality of food available to them. Furthermore, because the nutritional value of their main year-round food items (i.e. lichen and mosses) is relatively low [Project # 23. Please see Appendix 3-1] any reduction in the quantity and quality of caribou forage species could have a negative impact on caribou body condition and/or vital rates.
Preliminary evidence for a positive relationship between open, recently harvested areas and biting insect abundance indicates that recently harvested landscapes could expose caribou to higher levels of insect harassment [Project # 26. Please see Appendix 3-1]. Studies of the energetic consequences of insect harassment on other caribou subspecies and ecotypes have documented increases in costly avoidance responses (e.g. increased time spent standing or moving) and reductions in time spent feeding (Downes et al. 1985; Toupin et al. 1996). Preliminary results indicate that caribou may reduce time spent feeding when insect harassment is high, but there is no evidence for a marked increase in energetically costly behaviours [Project # 27. Please see Appendix 3-1].
The results of caribou diet analyses [Project # 22. Please see Appendix 3-1] and plant biomass and nutritional analysis [Project # 23. Please see Appendix 3-1] have enabled researchers to calculate energy available to caribou in different habitat types and seasons, which is a critical component of the caribou ecological energetics models [Project # 28. Please see Appendix 3-1]. Likewise, research focused on caribou energy costs [11-14] has provided insight into the impacts that caribou behaviour and environmental conditions have on energy levels, which are also a critical component of the ecological energetics models. The ecological energetics models will allow researchers to gain a better understanding of how habitat and diet may limit caribou populations and assess whether energetic balances for caribou are likely to differ between managed and unmanaged landscapes. The models will also provide detailed insights into the specific factors responsible for reducing net energy gain. Key results from all energy gain and energy loss studies (including direct data analysis and modelling efforts) are being integrated into a spatially explicit PVA (Section 3.2.13) for caribou that will allow researchers to explore the effects of energy balance on the long-term probability of caribou population persistence.
Finally, preliminary results from research on body condition and key vital rates [Project # 29. Please see Appendix 3-1] do not focus on identifying and assessing the impact of different mechanisms that underlie energy balance in caribou, yet they have allowed researchers to test some predictions associated with the Energetic Balance Hypothesis. Direct assessments of adult and calf body condition are somewhat consistent with predictions associated with the Energetic Balance Hypothesis, while results for vital rate-based assessments are less straightforward. Research into calf and adult body condition suggests that net energy gain may be lower in managed landscapes – which could have a direct or indirect impact on mortality rates in these areas (see Section 3.2.12). There is a trend towards higher pregnancy rates in the unmanaged landscape, but since this vital rate can be influenced by factors other than nutrition (see Brown et al. 2007 for a detailed discussion), the implications of this result for condition of adults in managed vs. unmanaged landscapes are not clear.
3.2.8 The sensory disturbance hypothesis: Human activity, caribou response and mitigation approaches
Research results suggest that the tendency of caribou to avoid roads became stronger as the amount of traffic increased.
Research into different techniques that can be used to either reduce traffic volume on roads or to help trees and plants grow on old roadbeds, indicates that some measures were more effective than others.
Research goals & objectives
To determine whether there is support for the Sensory Disturbance Hypothesis and to gain a better understanding of whether sensory disturbances from human activities affects caribou, researchers involved in the Research Program are studying the impacts of vehicle traffic, one of the major sources of sensory disturbance in the managed boreal forest. Researchers are also evaluating the effectiveness of different road decommissioning and rehabilitation techniques. This research can help inform planning for primary and resource access roads (CCP 3.9, 4.2.1, 7.2).
Methods & findings
How much vehicle traffic exists on road networks in managed and unmanaged landscapes?
To measure the volume of vehicle traffic moving through managed and unmanaged landscapes and determine how variable it is between seasons, MNRF researchers deployed electronic traffic counters on different road networks [Project # 30. Please see Appendix 3-1]. Data on the number of vehicles travelling along sampled roads were collected and used to model seasonal variation in vehicle traffic levels between managed and unmanaged landscapes. Preliminary results indicate that there was considerable variation in traffic volume between networks and between seasons (for all networks). Managed landscapes exhibited similar, relatively high traffic volumes in summer and low traffic volumes in winter (Figure 3-7). However, while Cochrane also exhibited relatively high traffic levels in spring, traffic levels were relatively low in Nakina. In contrast, traffic levels during fall were relatively high in Nakina and relatively low in Cochrane (Figure 10). Absolute traffic levels in the unmanaged landscape were well below those recorded in both the managed landscapes in all years/seasons. General traffic volume patterns in the unmanaged landscape also differed from those recorded in managed landscapes. Traffic levels in spring, summer and fall were relatively low, but peaked in winter (when traffic levels in managed landscapes were at or close to lowest levels) (Figure 3-7).
Figure 3-7. Comparisons of density of vehicle kilometres travelled (VKT) (VKT density = average daily kilometres travelled / land-based area km2) among the Research Program study sites from summer 2011 to spring 2013. The solid black line depicts seasonal variation in VKT density in Nakina (managed), the solid gray line depicts seasonal variation in traffic volume in VKT density in Cochrane (managed) and the dashed black line depicts seasonal variation in traffic volume in VKT density in Pickle Lake (unmanaged).
If you need alternate formats or communication supports to access this, please contact us.
Is caribou behaviour affected by variation in vehicle traffic on a major road?
Caribou telemetry data and traffic volume data was analyzed to determine whether traffic volume influenced caribou distribution and movement behaviour [Project # 31. Please see Appendix 3-1]. Analyses focused on evaluating whether caribou reactions to a major road (i.e. minimum distances, road crossing rates) changed in response to variation in traffic volume levels. Preliminary results indicated that the proximity of caribou to the road varied in association with changes in traffic volume – exhibiting an abrupt increase (i.e. a threshold-type response) as traffic volume increased above 65 vehicle counts per day or less (Figure 3-8). Preliminary results also indicated that caribou crossings of the major road were significantly lower than expected and occurred primarily during seasons when caribou are most mobile (i.e. fall and spring – Ferguson and Elkie 2004). Additionally, crossings of road segments with relatively high traffic levels occurred during periods when traffic volumes were low.
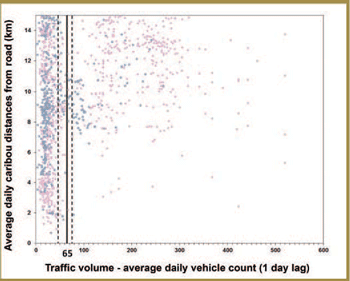
Figure 3-8. The relationship between traffic volume and average daily caribou distances to a major road. The blue dots represent caribou distances relative to the road in winter. The pink dots represent caribou distances relative to the road in summer. The solid black line represents the threshold traffic volume value (65 vehicle counts per day) at which minimum distances to the road increase and the dashed black lines demarcate the range of uncertainty (i.e. the 80% bootstrapped confidence interval) around the threshold estimate value.
If you need alternate formats or communication supports to access this, please contact us.
How effective are different approaches for restricting vehicle traffic on resource access roads?
To evaluate the effectiveness of different road decommissioning approaches, researchers collected electronic traffic counter data on single-lane resource access roads where different closure and decommissioning approaches had been applied [Project # 32. Please see Appendix 3-1]. Four different approaches were evaluated relative to each other and to open roads with no closure or deactivation measures: Roads closed year- round; seasonally-closed roads; deactivated roads; and roads that were both deactivated and closed. Human use during the moose hunting season (when recreational use of these roads is likely highest) was compared across approaches. Preliminary results indicated that seasonal road closures (i.e. two weeks or less during the gun hunting season for moose) led to the greatest reduction in traffic volume (predicted reduction = 92 %), while year-round closures were least effective (predicted reduction = 58%) (Figure 3-9). Deactivated roads and deactivated and closed roads were associated with intermediate levels of traffic reduction (Figure 3-9). Preliminary results also indicated that all approaches are less effective at reducing traffic volume on wider roads.
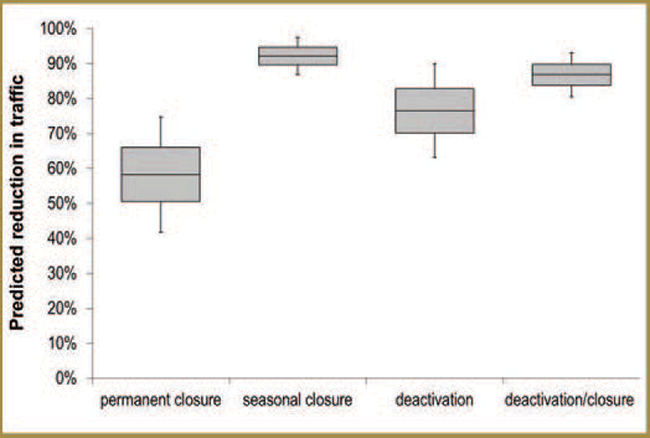
Figure 3-9. Estimated traffic reduction (%) on single lane resource roads in northern Ontario under different closure/deactivation treatments.
If you need alternate formats or communication supports to access this, please contact us.
How effective are different road reclamation techniques?
Researchers collected data on vegetation conditions (e.g. ground vegetation and shrub cover) and associated road and harvest conditions along several resource roads that had been subjected to different decommissioning and reclamation treatments [Project # 33. Please see Appendix 3-1]. A wide variety of treatments were applied and examined, including various combinations of full road re-contouring and reclamation, decommissioning, planting or seeding with desired species and natural abandonment.
Preliminary results indicated that reclamation treatments were more effective when combined with efforts to decommission the sampled road, likely due to reduced human use and protection of regeneration from vehicle damage. Additional preliminary results follow. Site preparation was more effective when combined with planting and seeding. Jack pine tended to grow better than black spruce when planted/seeded on roads, although high densities of seeds/seedlings may be needed to counter tree mortality after establishment. High amounts of gravel and high levels of compaction had negative impacts on vegetation establishment and growth and thus, regeneration success was better on winter roads.
Interpretation of research results
Research into the relationship between traffic volume and caribou behaviour [Project # 31. Please see Appendix 3-1] could not have been completed without the traffic volume research [Project # 30. Please see Appendix 3-1] described in this section. The results of this research can help resource managers separate and evaluate the different impacts roads can have on caribou populations (e.g. sensory disturbance vs. predator road use – see Section 3.2.10). Improved understandings of potential road impacts can help inform planning for primary resource access roads (MNR 2009 Action 4.2.1, 7.2).
Preliminary results [Project # 31. Please see Appendix 3-1] support some aspects of the Sensory Disturbance Hypothesis. They also provide insight into the potential consequences of constructing a high traffic volume road in the Continuous and Discontinuous Distribution. One potential impact of reduced caribou proximity to busy roads is avoidance of otherwise suitable habitat if it is adjacent to roads with moderate to high traffic volumes – a situation that might produce an effective reduction in the amount of habitat available for caribou. Additionally, if caribou avoid crossing busier roads, this could lead to increased fragmentation or isolation of local populations. Reduced crossings could also impede caribou re-occupancy of regenerating habitat.
Preliminary results of research on decommissioning of resource access roads [Project # 32. Please see Appendix 3-1], suggest that seasonal road closures may produce the greatest reduction in non-industrial traffic volume and consequently, help limit sensory disturbance impacts on caribou. However, while road closures may be effective at reducing traffic-related sensory disturbances, they may be less effective for reducing other negative impacts that roads can have on caribou (see Section 3.2.10 for a discussion of predator road use and enhanced predation risk).
Preliminary results from road reclamation research [Project # 33. Please see Appendix 3-1] support the use of the road-shed concept when planning and developing road networks, as construction methods and road use patterns that occur during the industrial lifespan of a given road, will affect the costs and success of future road reclamation efforts. A road-shed includes all roads that originate from a single point, typically a primary or secondary road, and in many ways is akin to a watershed (i.e. road network arrangement parallels connected streams and river arrangements within a watershed). When an area is first accessed for timber harvesting, the primary road is constructed first, and secondary and tertiary roads branch off of it. Road- shed based planning and development may help minimize the impacts of roads on caribou, as well as help maximize the effectiveness and minimize the costs of road reclamation efforts. Other measures that can improve the effectiveness of reclamation efforts include minimizing gravel additions and compaction while the roads are still in use, applying site preparation and seeding or planting of conifers at high densities, and favouring seeding/planting of jack pine over black spruce when site conditions are appropriate.
3.2.9 The apparent competition hypothesis: caribou relationships with predators and prey
Research indicates that in landscapes with considerable amounts of human disturbance, the abundance of moose and wolf densities were higher and the territories that wolves live in were smaller.
Research results suggest that in the three study areas investigated, moose were the primary year-round food source for wolves. Wolves tended to spend more time in forest types where there would likely be more moose (like mixedwood forests, which have considerable amounts of both deciduous and coniferous trees), and in areas where there were more resource access roads.
While research results indicate that caribou were not the main food source for wolves, predation by wolves was the most common cause of death in adult caribou.
Research goals & objectives
The Apparent Competition Hypothesis states that managed forests have characteristics (e.g. high forage availability) that attract and sustain high densities of alternate prey species (e.g. moose) which in turn, lead to higher densities of shared predators (e.g. wolves and bears) and higher incidental predation of caribou. The major predictions of the Apparent Competition Hypothesis include;
- Niche separation (i.e. separation of spatial distribution, habitat use and daily activities) between caribou and primary alternate prey (i.e. moose) and predators (i.e. wolves, bears).
- Niche overlap between moose and wolves.
- Higher densities of moose and wolves in managed landscapes.
- Dominance of moose or other prey/resources as predator food items.
- Higher predation-related mortality rates amongst caribou in managed landscapes.
Researchers involved with the Research Program and the Far North Caribou Project have been involved in several projects that test the predictions associated with the Apparent Competition Hypothesis and other predation-related hypotheses (e.g. Predator Road Use and Prey Escape). The results of these research projects can also be used to increase understanding of caribou mortality causes (MNR 2009 - Action 5.2), help assess the relationship between caribou and moose density (MNR 2009 - Action 5.4), and inform a habitat-based approach for managing prey and predator populations (MNR 2009 - Action 5.5).
Some of the projects described in this section are focused primarily on evaluating support for the Apparent Competition Hypothesis, but others also have implications for other predation-related hypotheses (i.e. the Apparent Competition, the Predator Road Use and the Prey Escape hypotheses – Section 3.2.1). Relevant results from this latter group of projects are also discussed in Sections that focus on the Predator Road Use Hypothesis (Section 3.2.10) and the Prey Escape Hypothesis (Section 3.2.11).
As mentioned above, a major prediction of the Apparent Competition Hypothesis focuses on predation-related mortality rates. The relative importance of predation events and other causes of caribou death have implications for all alternative hypotheses about factors that affect caribou persistence (see Section 3.2.1). For this reason, a more detailed description of research into caribou mortality causes and the factors affecting caribou survival rates appears in Section 3.2.12.
Methods & findings
How much overlap exists between habitat associations and spatial distributions of caribou, moose and wolves in winter?
To determine the general relationship between winter occupancy patterns of caribou, moose and wolves in the Far North of Ontario, the probabilities of occupancy for each species (see project [Project # 7. Please see Appendix 3-1] and Part 2, Section 2.5) were compared by evaluating their respective responses to the same general landscape attributes, including open water, terrain ruggedness, burned and harvested areas, wetlands and distance to human settlements [Project # 34. Please see Appendix 3-1]. Researchers also assessed the degree of spatial overlap between areas with high and low probabilities of occupancy for caribou and areas with high and low probabilities of occupancy for moose and wolves.
Caribou exhibited higher probabilities of occupancy in areas dominated by conifer, treed bog and sparse forest cover and avoided disturbed and regenerating areas (see Section 3.2.4. and the Far North portion of Figure 2-13 in Part 2, Section 2.5). Moose occupancy was high when amounts of disturbed habitat and terrain ruggedness (associated with mixedwood forest cover) were high and low when bog cover increased. In contrast to caribou and moose, probability of wolf occupancy had no significant relationship with coarse landscape attributes.
When occupancy modelling results were used to map the spatial distribution of all three species [Project # 34. Please see Appendix 3-1], the degree of spatial segregation between caribou and moose was considerable (Figure 3-10). Areas with high moose occupancy were found mainly in the more disturbed Ontario Shield Ecozone, while the area with highest caribou occupancy occurred along the transition between the fire- disturbed forests of the Ontario Shield Ecozone and the peatland complexes of the Hudson Bay Lowlands Ecozone (Section 3.2.4 and Figure 3-10). In contrast to the results for caribou and moose, spatial patterns in wolf occupancy were not as strong. Despite the lack of strong associations with landscape attributes, areas with high probabilities of wolf occupancy corresponded closely with high occupancy areas for both prey species (i.e. moose and caribou) (Figure 3-10).
What attributes are associated with higher probabilities of wolf use at the landscape level?
Researchers analyzed locations from GPS-collared wolves and caribou, aerial survey data for moose, and geo-spatial data for different landscape attributes (e.g. forest types, water bodies, shorelines and roads) to determine the relationship between wolf use and prey distributions, prey habitat, and features that might affect the ease of wolf movement through the landscape [Project # 35. Please see Appendix 3-1]. Preliminary results indicated that wolf density and space use were both concentrated near habitats preferred by moose, while there was much less overlap between caribou and either of the other species. In managed and unmanaged landscapes, spatial distributions of wolves and moose were similar and some community types favoured by moose (i.e. deciduous and mixed forests) were more strongly associated with wolf use than actual moose density was. Wolves also selected areas close to shorelines and dumpsites. Wolves in managed landscapes also appeared to select disturbed forests, while those in unmanaged landscapes didn’t. Preliminary results also suggested that caribou locations were negatively associated with areas with high wolf and moose use (particularly in managed landscapes and during winter), indicating partial spatial segregation of caribou from both wolves and moose.
Figure 3-10. Derived probability of occupancy of caribou, moose and wolves in the Far North of Ontario (adapted from Poley et al. 2013). The dashed line divides the Ontario Sheild Ecozone (west) and Hudson Bay Lowlands Ecozone (east). The caribou and moose models were derived in each ecozone spereately; the wolf model was derived across the whole study area.
If you need alternate formats or communication supports to access this, please contact us.
What attributes are associated with use, territory size and density of different wolf packs?
To determine what factors influence wolf habitat use, pack territory size and wolf pack density, researchers analyzed locations from GPS-collared wolves, aerial survey data for moose and geo-spatial data for different landscape attributes to determine the relationship between the habitat selection, space use patterns and abundance of different wolf packs, and the spatial distribution of roads, forest cover, water, topography and variation in moose density [Project # 36. Please see Appendix 3-1]. Preliminary results indicate that habitats selected by wolves at the pack level were similar to landscape level results [Project # 35. Please see Appendix 3-1]. Selection patterns did differ between packs, but wolves generally selected areas that were sloped and close to water and avoided areas with dense conifer cover. Local moose density was not a consistent predictor of pack use, but wolves did select forest types that are generally associated with high moose densities (i.e. deciduous and regenerating forests). Preliminary results from analysis of pack home ranges indicated that territory size decreased as moose density increased; consequently, there were more territories and packs in a managed landscape (which had higher moose densities), than there were in an unmanaged landscape (Figure 3-11).
Figure 3-11. GPS locations of collared wolves from different packs in Pickle Lake (unmanaged landscape) and Nakina (managed landscape). Different colours are used for different individuals (i.e., pack representatives) and some packs had more than 1 collared individual over the course of the study.
If you need alternate formats or communication supports to access this, please contact us.
How much overlap exists between daily activity levels of caribou, moose and wolves?
Researchers analyzed locations and accelerometer-based activity data from GPS-collared wolves, caribou and moose to determine whether the three species were active at the same times of day [Project # 37. Please see Appendix 3-1]. Moose data was obtained from a historic moose telemetry dataset. Preliminary results indicate that moose and wolves are active at similar times throughout the day and night, while there is lower correspondence between active periods for both species and those of caribou. Preliminary results from associated modelling work also suggest that these activity patterns could have appreciable effects on predation risk. Specifically, the lack of overlap between daily periods of high activity for caribou and those of wolves and moose may serve to reduce their risk of predation by wolves to some degree.
What do wolves eat and how important are caribou as a prey item?
Researchers used several different approaches to characterize activity data from GPS-collared wolves, caribou and moose wolf diets in managed and unmanaged landscapes to to determine whether the three species were active at the determine the relative importance of caribou as a prey item same times of day [Project # 37. Please see Appendix 3-1]. Moose data was obtained from a for wolves. Multiple approaches were used to compensate historic moose telemetry dataset. Preliminary results indicate for the respective advantages and disadvantages of different that moose and wolves are active at similar times throughout methods, which included possible biases towards detection of the day and night, while there is lower correspondence certain prey types and the restriction of inferences to certain between active periods for both species and those of caribou. Preliminary results from associated modelling work also suggest that these activity patterns could have appreciable effects on predation risk. Specifically, the lack of overlap between daily periods of high activity for caribou and those of wolves and moose may serve to reduce their risk of predation by wolves to some degree. times of the year.
- Winter wolf diets: Kill site investigations
Researchers analyzed locations and activity data collected from GPS-collared wolves during winter to identify and investigate locations where wolves killed and ate their prey (i.e. kill sites) [Project # 38. Please see Appendix 3-1]. Preliminary results indicate that moose was the dominant winter prey item in managed and unmanaged landscapes. Of the 232 kill sites investigated during winter, 95% were moose kills and only 3% were caribou kills. In the unmanaged landscape, two studied packs had territories with open-pit dumpsites and while these packs had large territories that they patrolled and defended, preliminary results from kill site investigations indicated that these wolves relied heavily on the dumpsites for feeding.
- Winter wolf diets: Stable isotope analysis of blood samples
To address concerns that kill site detection methods [Project # 38. Please see Appendix 3-1] might be biased towards large prey that take a long time to eat (e.g. moose), researchers used a specialized technique (i.e. stable isotope analysis) to analyze blood samples collected from wolves captured during winter [Project # 39. Please see Appendix 3-1]. Preliminary results suggest that caribou may make up a greater portion of winter wolf diets than indicated by kill site investigation results. The difference between the two methods may be due to underestimation of the importance of smaller prey associated with the kill site method [Project # 38. Please see Appendix 3-1], but preliminary results from stable isotope analyses also indicated that moose were still the dominant winter prey item for wolves in managed and unmanaged landscapes. Further refinement and confirmation of stable isotope analyses is currently underway.
- Late spring/early summer wolf diets: Scat analysis
To characterize wolf diets during the caribou calving and early post-calving periods, researchers analyzed wolf scats collected from May-June [Project # 40. Please see Appendix 3-1]. Preliminary results indicated that moose were the dominant prey item (42% of scat content volume) during and immediately after caribou Beaver were also an important food source (34% of scat content volume), while caribou calf remains were only identified in a single scat.
- Spring-autumn wolf diets: Stable isotope analysis of hair samples
To characterize wolf diets from spring to autumn, researchers analyzed wolf hair samples collected during capture and collaring efforts [Project # 41. Please see Appendix 3-1]. Preliminary results indicate that the importance of caribou in wolf diets increased from relatively low levels in spring and early summer to higher levels in late summer, but caribou were still a much less important food item than moose (which made up just over half of late summer wolf diets), snowshoe hare and beaver. Analysis of wolf hair samples is still ongoing.
What do bears eat in late spring early summer and how important are caribou as a prey item?
To determine the relative importance of caribou as a prey item for bears in late spring/early summer researchers characterized the contents of bear scats collected from late May to late June [Project # 42. Please see Appendix 3-1], which spans the period when caribou calves would have been most vulnerable to bear predation (Pinard et al. 2012). Over 600 bear scats were collected over two summers. Preliminary results indicate that the vast majority of the total scat biomass (i.e. over 90%) was comprised of grasses, sedges, mosses or fruit. Only two scats (i.e. <0.01%) contained caribou calf remains and most remaining scat volume was comprised of insects, moose, beaver, snowshoe hare, birds and small mammals.
Interpretation of research results
Obtaining an improved understanding of the relationship between caribou and their predators and clarifying how these relationships might be altered in managed landscapes is important for identifying the specific factors that might be driving caribou declines and informing development of best management practices for minimizing predator impacts in managed areas.
The results presented here provide support for the major predictions of the Apparent Competition Hypothesis.
Research conducted at coarse and fine spatial scales suggests that there is a high degree of overlap between the niches of moose and wolves (i.e. daily activity patterns, habitat use and spatial distribution), but appreciable separation between the preferred niches of moose and caribou. The amount of favourable moose habitat and moose densities are both higher in managed areas, while wolf pack territories are smaller and more densely distributed.
All preliminary results from analyses of wolf diets [38-41] indicated that moose are the most important year-round prey item, while caribou are secondary or even tertiary prey, with relative importance that changes seasonally. The implications of the black bear diet results [Project # 42. Please see Appendix 3-1] for the Apparent Competition Hypothesis are less clear, as bear diet analyses are much more limited than those carried out for wolves. However, while there is a small amount of evidence for bear predation on caribou (also see Section 3.2.12) this evidence indicates that caribou likely represent only a minor component of bear diets.
From a caribou conservation perspective, the results described here provide evidence that supports minimizing the positive impact that harvesting practices can have on the abundance of alternate prey and predators. Practices that promote the regeneration of naturally occurring conifer cover and avoid producing mixedwood and deciduous forest cover in amounts that exceed those expected to occur under a natural disturbance regime, may help achieve a reduced abundance of alternate prey and predators (see the Boreal Landscape Guide (OMNR 2014) and Section 3.2.5). At the stand and site level, this could involve the application of relatively intensive silvicultural treatments: mechanical site preparation, planting or aerial seeding of conifers, herbicide applications (although see Section 3.2.6 for discussion of the negative impacts of herbicides on caribou forage) and prescribed burning.
Finally, wolf habitat selection [Project # 35. Please see Appendix 3-1] and diet analyses [Project # 38. Please see Appendix 3-1] suggest that garbage disposal sites associated with human settlements and activities can serve as a major food source for wolves. Just as higher densities of alternate prey can support higher densities of predators it appears that human wastes might support higher densities of wolves. While caribou may not be the primary food source for packs that target dumpsites, the two packs that fed primarily at dumpsites still had large territories and accordingly, still kill live prey when they encounter them. These results provide support for minimizing the number of dumpsites and can help inform the location of new dumpsites.
3.2.10 The predator road use hypothesis: enhanced hunting efficiency
Research suggests that the use of resource access roads by wolves allowed them to move more quickly throughout their territories and improved their hunting success.Research results indicate that if caribou use features on the landscape that are also selected by wolves and which help wolves to be more efficient at hunting moose (like access roads and mixedwood forests), they may be more at risk of being detected and killed by wolves.
Research results indicate that if caribou use features on the landscape that are also selected by wolves and which help wolves to be more efficient at hunting moose (like access roads and mixedwood forests), they may be more at risk of being detected and killed by wolves.
Research goals & objectives
The Predator Road Use Hypothesis states that when available, wolves use roads to explore their territories and this road use makes them more efficient at hunting and increases the threat they pose to caribou. As part of the Research Program, researchers have initiated several projects to assess the amount of support for this hypothesis (and in some cases, other predation-related hypotheses). This research can also help to improve understandings of caribou mortality causes (MNR 2009 - Action 5.2), predator impacts on caribou (MNR 2009 - Action 5.5), and potential impacts of roads on caribou and caribou habitat (Action 3.7.3, 3.9, 7.2).
Methods & findings
Does road use influence the hunting efficiency of wolves?
Researchers used location data from GPS-collared wolves and associated kill site investigation data [Project # 38. Please see Appendix 3-1] to determine whether time between wolf kills (i.e. hunting efficiency) was affected by landscape attributes (e.g. roads), moose density, weather and/or pack size [Project # 43. Please see Appendix 3-1]. Research efforts were limited to managed landscapes due to the reliance on dumpsite scavenging documented for multiple packs in the unmanaged landscape [Project # 38. Please see Appendix 3-1]. Preliminary results for managed landscapes indicate that distance from roads and speed of wolf movements had the greatest influence on hunting efficiency. The chance of making a successful kill increased when wolves were closer to roads and when their travel speeds were high. Higher snow depths and increases in the amount of mixedwood cover (i.e. preferred moose habitat) also led to improved hunting efficiency, but neither of these features were as influential as roads. Preliminary results indicated that several factors that were expected to affect wolf search efficiency had little detectable influence on hunting efficiency: Prey density, weather conditions, pack size and landscape attributes expected to enhance prey detection (i.e. cutblocks, regenerating forests and topography).
Are roads associated with moose kill sites?
Researchers analyzed landscape attribute data, environmental condition data and wolf and moose density data associated with locations where wolves killed their dominant winter prey (i.e. moose kill sites [Project # 38. Please see Appendix 3-1]), to determine the influence that habitat type, predator density, prey use and prey vulnerability have on the probability that a wolf will make a kill [Project # 44. Please see Appendix 3-1]. Preliminary results indicated that wolves tended to kill moose in areas located away from open water, lowland and human settlements, but close to secondary and tertiary roads (e.g. resource access roads). In addition to road effects, preliminary results indicated that the probability of killing a moose increased in areas with either high moose use or high wolf density, but in areas where both are high, this effect wasn’t as strong. At high wolf densities, the probability of killing a moose also increased sharply in deep snow conditions, but snow depth had little or even negative effect on hunting success when wolf use and snow depth were both low.
Do wolves select roads at landscape and/or pack scales?
The methods used to determine what factors influence wolf habitat use at landscape [Project # 35. Please see Appendix 3-1] and pack scales [Project # 36. Please see Appendix 3-1] are discussed in detail in Section 3.2.9. Preliminary results from both sets of analyses were consistent with wolf selection for roads, especially in summer. At the landscape scale, areas close to rivers and edges of large lakes were also selected, but settlements were avoided [Project # 35. Please see Appendix 3-1]. At the pack scale, selection for roads was strongest amongst packs with territories that contained large road networks [Project # 36. Please see Appendix 3-1]. Wolf responses to forest types and variation in prey density are discussed in Section 3.2.8.
Interpretation of research results
The results presented here provide support for the Predator Road Use Hypothesis, insofar as it applies to wolves - the primary predator of adult caribou (see Section 3.2.12). Specifically, wolves appear to select roads at multiple scales [Project # 35, 36. Please see Appendix 3-1] and preliminary results suggest that the use of roads by wolves appears to decrease time between kills [Project # 43. Please see Appendix 3-1] and increase the probability of making a successful moose kill [Project # 44. Please see Appendix 3-1]. Consequently, using or travelling through areas with extensive roads networks likely increases predation risk for caribou. While caribou may avoid busier roads with high levels of associated sensory disturbance, they may not respond as strongly to resource roads with lower traffi volumes (see Section 3.2.8). Thus, they may be more vulnerable to predation in areas with extensive low-traffi volume road networks.
If caribou represent secondary or tertiary prey for wolves (see Section 3.2.9) and they are generally killed following opportunistic encounters, then it is likely that the landscape attributes associated with higher wolf use and greater hunting success for primary prey (i.e. resource access roads and mixedwood cover) will be associated with higher levels of predation risk for caribou. Thus these results can help inform road planning that could minimize the infl of road networks on wolf hunting success and predation risk for caribou. One measure that might reduce the suitability of existing roads as a travel corridor for wolves could involve restoring old road beds to pre-development forest conditions. Implementing effective road closure approaches (e.g. seasonal road closures) may reduce vehicle traffic but research indicates that efforts to regenerate vegetation on roads (in an effort to reduce their value as movement corridors for predators) may require more active reclamation measures (see Section 3.2.8).
3.2.11 The prey escape hypothesis: Restricted space use and increased detectability
Research goals & objectives
Research suggests that in winter, when individual caribou were close to recent disturbance and road developments they were more likely to return to areas that they had used in previous winters.
Research results indicate that wolves spent more time and were more successful at capturing prey in forest types selected by moose (such as mixedwood forests), while they avoided or were indifferent to forest types that were preferred by caribou (such as coniferous forests) and these forest types did not improve their hunting success.
The Prey Escape Hypothesis states that human disturbances and developments cause caribou in managed landscapes to restrict their movements to remaining patches of mature forest, which makes them more readily detectable by predators and more vulnerable to predation. Several research projects undertaken as part of the Research Program, and the monitoring and assessment activities (see Part 2 for more details) including the Far North Caribou Project and Integrated Range Assessments have assessed the degree of support for different aspects of this hypothesis. The results of these projects will increase the understanding of habitat- based approaches to manage predator populations (MNR 2009 Action 5.5).
Methods & findings
Are caribou home ranges sizes influenced by human disturbance?
The methods used to study factors that influence caribou home range size [Project # 14. Please see Appendix 3-1] are described in detail in Section 3.2.4. Results indicated that annual home ranges used by individual forest-tundra and forest-dwelling woodland caribou were larger when the abundance of suitable caribou habitat (i.e. conifer forest and treed wetlands) was low and smaller when the amount of suitable caribou habitat was high.
Is caribou fidelity to seasonal ranges influenced by human disturbance?
The approach used to study the factors that affect caribou fidelity to seasonal ranges [Project # 12. Please see Appendix 3-1] is discussed in detail in Section 3.2.4. One key preliminary result was relevant to the Prey Escape Hypothesis. Namely, that traditional use of specific winter sites increased when these sites were closer to roads and young forests and decreased when these sites were closer to mature upland forests. In other words, individuals were more likely to return to the same wintering location when they lived in areas with high amounts of disturbance and development and less likely to use the same location over multiple winters when surrounded by the sort of mature forests that are more abundant in unmanaged landscapes.
Is wolf habitat selection influenced by caribou density or proximity to suitable caribou habitat at landscape and/or pack scales?
The second major prediction of the Prey Escape Hypothesis are discussed in detail in Section 3.2.9. Two preliminary is that if caribou are restricted to fewer and smaller areas of results from this research were relevant to the Prey Escape suitable habitat in managed landscapes, wolves will be better Hypothesis. The first is the positive influence that forest able to detect and target caribou in these areas. However, types commonly selected by moose (e.g. deciduous, mixed results from research into wolf hunting success [Project # 43. Please see Appendix 3-1] and and regenerating forests) have on wolf use. The second is habitat selection at landscape and pack scale [Project # 35, 36. Please see Appendix 3-1] were wolf avoidance of forest types commonly selected by caribou not consistent with these predictions. While wolves selected (i.e. conifer forests).
Is wolf hunting success influenced by caribou density or proximity to suitable caribou habitat?
The approach used to determine what factors influence wolf hunting success [Project # 43. Please see Appendix 3-1] is described in detail in Section 3.2.10. Key preliminary results from this research are relevant to the Prey Escape Hypothesis: the improved hunting success that wolves experienced in suitable moose habitat (i.e., mixedwood forests), combined with the finding that use of conifer-dominated forest types had no impact on how successful wolves were at killing prey.
3.2.12 Caribou mortality factors: Evaluating support for multiple alternative hypotheses
Research indicates that the most common cause of death amongst adult caribou in both managed and unmanaged landscapes was predation, and predation-related and overall mortality rates were highest in the landscape with the highest levels of human disturbance.
Research suggests that the relationship between different landscape features (such as different forest types and roads) and caribou mortality levels (predation-related and overall) is still being studied.
Research into the causes of adult caribou deaths did not document any examples where caribou deaths were caused by humans. These results suggest that the effects that human activities and developments can have on the landscape may have a greater impact on caribou than direct, human-caused mortalities.
Preliminary research results suggest that adult caribou living in areas with considerable amounts of human disturbance (like harvesting and roads) may be in poorer physical condition than those living in unmanaged landscapes. However, this trend did not appear to result in greater numbers of deaths from condition-related causes amongst adult caribou living in these landscapes.
Interpretation of research results
The results from research conducted to date do not provide much support for the Prey Escape Hypothesis. One major prediction of this hypothesis is that if forest management and development activities reduce the total amount and/mortality or change the arrangement of suitable caribou habitat (e.g. mature conifer forests), caribou will restrict their movements to remaining habitat patches. Research into the factors that influence caribou home range sizes [Project # 14. Please see Appendix 3-1] documented the opposite results - rather than restricting death was detected to determine cause of death [Project # 45. Please see Appendix 3-1]. Caribou mortality data were used to characterize seasonal patterns in cause of death and compare overall and cause-specific mortality rates between managed and unmanaged landscapes.
A total of 48 adult female caribou mortalities were documented and investigated. Preliminary results suggest that wolf predation was the most common direct cause of death in managed and unmanaged landscapes. Other direct causes included bear predation (three caribou), pregnancy- related complications, injury and drowning, while old age may have been a contributing factor in a few cases. None of the investigated mortalities appeared to be caused by humans.
Research goals & objectives
The alternative hypotheses evaluated as part of the Research Project (e.g. Energetic Balance, Disturbance, Apparent Competition, Predator Road Use and Prey Escape - outlined in Section 3.2 1) make different predictions regarding the cause of caribou deaths and/or the factors that influence caribou mortality rates. To determine which factors have the greatest influence on caribou survival, researchers investigated causes of adult caribou deaths and studied the influence of different factors on caribou mortality rates.
Researchers also analyzed more limited evidence from video collar data to improve understandings of the factors that influence calf survival. Additionally, while disease is not believed to have major impacts on caribou populations in Ontario, there have been some concerns about potential impacts on caribou health, which could affect their vulnerability to other causes of mortality. To address these concerns, researchers studied incidence levels of diseases that might have a negative effect on caribou condition.
The results of these research projects can provide information related to human caused caribou mortalities (MNR 2009 - Action 5.2). While research results are limited to documenting mortality information for collared caribou, they provide some insight into the relative importance of different causes of caribou death and the factors that influence caribou mortality rates.
Methods & findings
What factors influence adult caribou mortality rates and causes of death?
Researchers monitored location and activity data from GPS-collared caribou to detect mortalities and conducted field investigations of mortality sites as soon as possible after a Preliminary results also suggested that while annual survival rates in all landscapes were relatively high (i.e. >75%), mortality rates were higher in the most heavily disturbed managed landscape. This difference appears to be driven by higher predation-related mortality rates, while rates of mortality due to other causes were similar across managed and unmanaged landscapes. Mortality risk also varied between seasons. Preliminary results indicate that the risk of dying from predation was lowest during seasons when caribou were relatively sedentary (i.e. late spring through summer) and peaked during seasons where caribou were more mobile (i.e. autumn/early winter and early spring). The risk of dying from other causes was also lowest during late spring/early summer (i.e. the calving and early nursery period), but it peaked in late summer. Additional analyses are currently underway to determine whether specific landscape attributes (e.g. linear features, disturbance and habitat) influence adult caribou mortality risk (see Appendix 3-4).
Does condition of adult caribou at death differ between managed and unmanaged landscapes?
When available, intact leg and jaw bones were collected from caribou carcasses at investigated mortality sites [Project # 45. Please see Appendix 3-1] and researchers analyzed the amount and appearance of bone marrow fat to characterize condition at death and determine whether it differed between managed and unmanaged landscapes [Project # 46. Please see Appendix 3-1]. Damage from predators, scavengers and decomposition limited the number of bone marrow fat samples that could be analyzed.
For the caribou that were evaluated, there were no general differences in the condition of animals that died in managed and unmanaged landscapes and no general differences between caribou that died from predation and those that died from other causes. However, when non-predation- related deaths were analyzed separately, bone marrow fat content was lower among caribou that died in an unmanaged landscape than it was amongst animals that died in managed landscapes, suggesting that animals dying from causes other than predation in the unmanaged landscape may have been in poorer condition when they died.
Does caribou calf condition at death differ between managed and unmanaged landscapes?
Data on the timing and cause of caribou calf deaths was very limited. Researchers analyzed very high resolution video data from a small sample of GPS-collared (video) females to identify calf births and determine when they appear to have died [Project # 29. Please see Appendix 3-1]. There was no direct video record for most calf deaths, so for GPS-collared (video) cows that gave birth to live calves, the start of a continuous, sustained period with no calf sightings was treated as evidence that a calf had died. Visual evidence of calf body condition was also used to characterize calf condition at birth and prior to death.
In general, small sample sizes of GPS-collared (video) caribou prevent strong conclusions from being drawn about differences in calf survival rates, calf condition and timing of calf deaths between managed and unmanaged landscapes. However, some general patterns were apparent (also see Section 3.2.7). Preliminary results indicated that most viable calves born in all study areas had died by early July (i.e. before two months of age). Preliminary results from calf condition assessments, suggest that calves in managed landscapes may be in poorer condition prior to death than calves born in unmanaged landscapes, but further analyses are needed to confirm this trend.
Disease/infection
To characterize the diversity and incidence levels of members of a particular family of roundworm parasites (Protostrongylidae) amongst adult caribou living in managed and unmanaged landscapes, researchers analyzed fecal samples collected when caribou were captured and collared [Project # 47. Please see Appendix 3-1]. When possible, parasites were identified to the species level using DNA methods. Preliminary results indicate that infection levels were relatively low amongst caribou living in an unmanaged landscape (i.e. 13%) and relatively high (22% to 23%) amongst caribou living in managed landscapes. While it was only possible to identify which Protostrongylid parasites species were present for a fraction of analyzed samples, preliminary results yielded evidence for infection by two different parasites in all studied landscapes: an undescribed species of Varestrongylus (a lungworm) and Parelaphostrongylus andersoni (a muscleworm).
Interpretation of research results
Information regarding caribou mortality levels and causes of adult and calf deaths can improve our understanding of the factors that drive caribou population dynamics and influence the probability that caribou will persist over the long-term (MNR 2009 – Action 5.2). Preliminary results indicate that the most common cause of death amongst adult caribou in managed and unmanaged landscapes is predation [Project # 45. Please see Appendix 3-1]. Research indicates that this is also true for caribou calves living in other jurisdictions (e.g. B.C. and Québec – Gustine et al. 2006, Pinard et al. 2012), but this has yet to be demonstrated for Ontario (see Section 3.4). Relatively high levels of predation-related mortality documented in the most intensively managed landscape is consistent with predictions of all predation-related hypotheses (i.e. Apparent Competition, Predator Road Use and Prey Escape Hypothesis - Sections 3.2.9, 3.2.10 and 3.2.11, respectively).
Results also indicated that the risk of predation varied seasonally: mortality rates were lowest from late spring to early summer, while overall and predation-related mortality rates were relatively high from late fall to early spring. These preliminary patterns appear to differ somewhat from seasonal mortality patterns documented in other jurisdictions, where highest general and predation-related mortality rates occurred in summer (e.g. Wittmer et al. 2005; Whittington et al. 2011; Rettie & Messier 1998; Courtois et al. 2007).
Ongoing research into the impacts of different landscape attributes on caribou survival rates and probability of predation-related mortality (Appendix 3-4) will help gauge the relative degree of support for different predation-related hypotheses. Of the six hypotheses that attempt to explain the correlation between human disturbance and the population declines and/or range retractions of caribou (see Section 3.2.1), three focus on the effects of human developments and landscape alterations on the behaviour and population dynamics of predator species (see Sections 3.2.9, 3.2.10 and 3.2.11). All of the hypothesized impacts are predicted to result in increased predation rates on caribou and, to date, mortality data for adult caribou in Ontario have matched these predictions.
No human caused mortalities were documented as part of the research program; however, mortality data from studies of collared caribou could underestimate the rates of some types of human-caused mortalities. Other studies of VHF radio or GPS collared animals (e.g. woodland caribou, white-tailed deer, grizzly bears, Eurasian lynx and Scandinavian wolves) have identified several potential sources of bias that could include the following:
- Multiple caribou in a group may be killed as part of a single hunting event (Rudolph et al. 2012; CBC 2007); therefore, results based on the fate of randomly selected individuals could underestimate total harvest impacts (Rudolph et al. 2012).
- Hunters may attempt to cover up evidence of hunting mortalities of radio-collared animals by destroying or disposing of radio-collars (McLellan et al. 1999, Andrén et al. 2006; Liberg et al. 2012). Non-subsistence harvest of caribou has been illegal in Ontario since 1929 (Darby et al. 1986), thus concerns about legal penalties could lead poachers to engage in such activities to avoid detection (e.g. Andrén et al. 2006).
- Hunters may avoid harvesting collared animals (e.g. McLellan et al. 1999, Jacques et al. 2011), so human harvest rates amongst collared animals may underestimate total human harvest rates (Jacques et al. 2011).
- Train collisions can cause multiple deaths in one group when they occur (e.g. a single accident killed 12 caribou in Nakina during the winter of 1988/89 – Cumming and Beange 1993), however, if they are relatively infrequent, short-term studies of randomly selected caribou may fail to detect them.
While these potential sources of bias should be acknowledged, preliminary results from research conducted in managed and unmanaged landscapes (located at, or relatively close to the southern boundary of the Continuous Distribution), suggests that direct human impacts on caribou mortality rates might not be as important as indirect impacts of human disturbance on ecological communities (e.g. landscape alteration and consequent impacts on the abundance of alternative predators and prey).
The Energetic Balance Hypothesis predicts that caribou living in managed landscapes will be in poorer condition than caribou in unmanaged landscapes. While caribou that are highly nutritionally stressed could die directly from malnutrition, poor condition could also increase vulnerability to other direct causes of death (e.g. predation). Preliminary results indicate that adult caribou living in managed landscapes may be in poorer condition than those living in unmanaged landscapes (see Section 3.2.7), but to date, none of the preliminary results described in this Section [Project # 45, 46. Please see Appendix 3-1] suggest that this trend may be resulting in higher adult mortality rates.
No direct efforts to collect detailed data on caribou calves during their first few months of life have been made in Ontario. This is partially due to concerns about the negative impacts capture and collaring activities might have on young caribou during a vulnerable life stage. However, preliminary results from analyses of video data from GPS-collared cows with calves (described in this Section and Section 3.2.7) indicates that there may be a difference between the nutritional status of calves in managed and unmanaged landscapes. This preliminary trend needs to be confirmed with additional research.
Calf survival is a vital rate that varies substantially between caribou ranges (see Section 2.3.3.2) and a review of population dynamics in large herbivores suggests that it can be the main determinant of population state (Galliard et al. 1998). Thus, efforts to reduce the uncertainty associated with causes of calf deaths (including the potential role of calf condition as a contributing factor to early calf mortalities) are important. However, gaining greater understanding of the causes of caribou calf deaths in managed and unmanaged landscapes would require the collection of different data than that collected as part of current research activities (e.g. Pinard et al. 2012).
Finally, while disease is not believed to have a major influence on caribou populations in Ontario, the diversity of caribou parasites and their potential impacts on caribou health are not well-documented. Parasites from the Protostrongylid family can cause moderate to serious disease in infected caribou. The Protostrongylid of greatest concern is brain worm (Parelaphostrongylus tenuis), which is carried by white-tailed deer and can be fatal to moose and caribou. While preliminary analyses did not detect P. tenuis in sampled caribou [Project # 47. Please see Appendix 3-1], fecal analysis may not be the best detection method for this parasite. In situations where P. tenuis is a concern, a monitoring approach focused on sampling white-tailed deer in areas where deer and caribou overlap could be more effective.
Even in cases where disease doesn’t result in death, Protostrongylid infections among caribou could have negative health effects that increase their vulnerability to other mortality factors. The two Protostrongylid species identified in managed and unmanaged landscapes [Project # 47. Please see Appendix 3-1] both have potential to produce negative health effects in caribou. P. andersoni is a muscle-worm that causes muscular and pulmonary disease and Varestrongylus can cause mild pneumonia. As with brain worm, white-tailed deer are good hosts for both parasites, while prevalence and intensity of infection among moose is extremely low. Preliminary trends towards higher incidence of Protostrongylid infections amongst caribou living in managed landscapes may be related to higher densities of white-tailed deer in areas that have more suitable deer habitat (e.g. regenerating forests)
Methods & findings
Researchers developed a simple PVA model structure and applied it to caribou populations living in managed and unmanaged landscapes, by incorporating vital rate estimates obtained from monitoring GPS-collared caribou in the Research Program study areas [Project # 48. Please see Appendix 3-1].
The simple caribou PVA model has the following features:
- Incorporates population or range-specific estimates of adult survival, pregnancy, and calf
- Predicts the consequences of these estimates for intrinsic rates of population change (l).
- Can be used to evaluate the sensitivity of l to changes in vital rates and estimate the probability that a population collapse to low levels within a specified period of time (e.g. 100 years).
In addition to field data on caribou vital rates, the second, spatially-explicit PVA also uses a novel, researcher-developed simulation model of caribou movement that incorporates the key biological characteristics of caribou in Ontario [Project # 49. Please see Appendix 3-1]. To ensure caribou movement was represented appropriately in the model, researchers analyzed location data from GPS- collared caribou to determine how caribou movements
3.2.13 Caribou population viability analysis models that is more complex) that can be used as forest composition, developments and different resource
Research goals & objectives
One of the principal goals of the Research Program is to develop two models for caribou Population Viability Analysis (PVA). The first is a simple, single species model that uses only survival and recruitment estimates to predict the potential for long-term growth or decline of caribou in specific populations or ranges. The second is a highly detailed, spatially-explicit model that integrates the full array of factors that affect caribou populations, including forest structure, food availability, snow cover, predation risk and moose densities. The spatially-explicit caribou PVA model is designed to be applicable to any caribou range in Ontario. Both PVA models have been designed as tools to conduct assessments of the long-term probability of persistence (i.e. viability) of specific caribou populations.
These models can help researchers determine the relative influence of different factors on caribou population health. They can also be used as decision support tools to help:
- Identify benchmarks for population viability (MNR 2009 Action 1.1); and
- Inform the assessment of the potential impacts that different resource development proposals might have on caribou population persistence (MNR 2009 Action 3.7, 7.2).
Methods & findings
Researchers developed a simple PVA model structure and applied it to caribou populations living in managed and unmanaged landscapes, by incorporating vital rate estimates obtained from monitoring GPS-collared caribou in the Research Program study areas [Project # 48. Please see Appendix 3-1]. The spatially-explicit caribou PVA model has the following features:
- Identify benchmarks for population viability (MNR 2009 Action 1); and
- Inform the assessment of the potential impacts that different resource development proposals might have on caribou population persistence (MNR 2009 Action 7, 7.2).
The simple caribou PVA model has the following features:
- Incorporates population or range-specific estimates of adult survival, pregnancy, and calf recruitment.
- Predicts the consequences of these estimates for intrinsic rates of population change (λ).
- Can be used to evaluate the sensitivity of λ to changes in vital rates and estimate the probability that a population will collapse to low levels within a specified period of time (e.g. 100 years).
In addition to field data on caribou vital rates, the second, spatially-explicit PVA also uses a novel, researcher-developed simulation model of caribou movement that incorporates the key biological characteristics of caribou in Ontario [Project # 49. Please see Appendix 3-1]. To ensure caribou movement was represented appropriately in the model, researchers analyzed location data from GPS-collared caribou to determine how caribou movements respond to forage availability, wolf occurrence, moose occurrence, biting fly abundance, ambient temperature and snow-depth.
The spatially-explicit caribou PVA model has the following features:
- Incorporates key biological features of caribou - including individual movement, selection and avoidance of different landscape attributes and interactions with predators and alternate prey species.
- Can be tailored to model and assess the viability of any caribou population using basic data on geospatial (e.g. forest types, linear features) and biological (local moose and wolf densities) characteristics. In the absence of local biological data, province-wide estimates for key demographic variables are assumed.
- Can be used for status assessment, and to inform planning efforts.
Key components of the spatially-explicit caribou PVA model are listed in Appendix 3-5, along with data sources used to initialize the model. The approach used to model caribou movement [Project # 49. Please see Appendix 3-1] is a critical component of the spatially-explicit PVA tool [Project # 48. Please see Appendix 3-1], as it specifies how modelled caribou agents respond to and interact with their surrounding environment, which in turn, affects adult mortality and recruitment rates and consequently, population growth (Figure 3-12).
Figure 3-12. Simulated movement trajectory of a single caribou across part of the Nakina landscape, as part of the spatially-explicit caribou PVA model [Project # 48, 49. Please see Appendix 3-1]. The shading of hexagons reflects the degree of predation risk during the summer, with red being high risk, yellow being low, and blue being small water bodies. Predation risk values were derived from an analysis of habitat selection by GPS-collared wolves living in the same landscape.
If you need alternate formats or communication supports to access this, please contact us.
For each run of the model, changes in the size of a caribou population are simulated many times (e.g. 1000) over a user-specified time period (e.g. 100 years). At the end of this process, a long-term probability of population persistence can be obtained by summarizing across the results of all simulations. The spatially-explicit model can be applied to any range within the Continuous Distribution. However, simulations conducted to date are based on managed and unmanaged Research Program study sites (i.e. Nakina and Pickle Lake).
Preliminary results from the spatially-explicit PVA model [Project # 48. Please see Appendix 3-1] (which includes the caribou movement model [Project # 49. Please see Appendix 3-1]) suggest that caribou in the managed landscape may have a low probability of persisting over the next 100 years, due to reduced access to food and elevated predation risk associated with high mixedwood covers and extensive road networks (which supports a relatively high abundance of moose and wolves). In contrast, preliminary results for the unmanaged landscape suggest that caribou in this area have a low probability of exhibiting dramatic declines in abundance. These preliminary predictions are consistent with preliminary projections for the managed and unmanaged study areas generated by the simple caribou PVA model and thus, the application of both models suggests that current landscape patterns in the managed landscape (i.e. Nakina) may not be sufficient to enable caribou to persist in the long- term under current conditions.
Interpretation of research results
The PVA models can be used as decision support tools to help evaluate potential impacts of resource management and development activities on caribou populations (MNR 2009 Action 1.1, 3.7, 7.2). Results of research conducted to date suggest that multiple factors associated with human disturbance and development may be having a negative impact on caribou populations, which is consistent with the Cumulative Effects Hypothesis (Section 3.2.1). The spatial PVA approach described here enables the evaluation of the cumulative effects of these factors on caribou population status. It can also be used to help assess the impacts that different development or disturbance scenarios may have on the long-term population viability of caribou.
One of the major barriers that could limit the usefulness of a spatial PVA model relates to whether it’s accessible and easy to use. To overcome this barrier, several features have been developed to make the model accessible to a wide variety of users (e.g. movies of simulated caribou movements, visualizations of changes in population sizes over time).
Users are also able to control the initial conditions for a number of model variables, including caribou population size at the start of the simulations, habitat characteristics, timber harvesting levels, and abundances and harvest rates for moose and wolves. These features allow the PVA model to be used as a tool to evaluate different planning and development scenarios that can be compared and used to rank different options.
3.2.14 Caribou resource selection functions (RSFS)
Researchers developed seasonal Resource Selection Functions (RSFs) for several ranges within the Far North of Ontario, as well as two southern ranges. Results from the RSFs suggest that the tendencies of caribou to select or avoid particular landscape features (like different forest types and disturbance) differed between ranges and between seasons. However, some general tendencies were fairly consistent. For example, caribou consistently selected coniferous forest, treed and open peatlands and sparsely treed areas and avoided mixed and deciduous forests, roads and natural disturbance.
The RSF models can be used to help map areas of suitable caribou habitat across these ranges and within different seasons. They can also be used as support tools to help assess the amount and distribution of caribou that changes in forest composition, developments and different resource management activities may result in.
There is uncertainty regarding the potential impacts that increased human disturbance and climate change might have on habitat availability and long-term persistence of caribou that inhabit the Far North of Ontario. To contribute to efforts to integrate caribou habitat considerations into planning initiatives in this region (MNR 2009: Action 3.1), researchers developed Resource Selection Functions (RSF) for several caribou ranges within the Far North of Ontario, as well as two southern caribou ranges, in order to
- Identify landscape attributes associated with high and low probabilities of caribou use at the broader landscape scale.
- Map suitable caribou seasonal habitat within these ranges.
- Gain a better understanding of the factors that influence caribou habitat selection.
Methods & findings
Researchers developed seasonal Resource Selection Functions (RSFs) for several ranges within the Far North of Ontario, as well as two southern ranges. Results from the RSFs suggest that the tendencies of caribou to select or avoid particular landscape features (like different forest types and disturbance) differed between ranges and between seasons. However, some general tendencies were fairly consistent. For example, caribou consistently selected coniferous forest, treed and open peatlands and sparsely treed areas and avoided mixed and deciduous forests, roads and natural disturbance.
The RSF models can be used to help map areas of suitable caribou habitat across these ranges and within different seasons. They can also be used as support tools to help assess the amount and distribution of caribou that changes in forest composition, developments and different resource management activities may result in.
To complete the RSF analysis, researchers worked with caribou GPS-collar data that was collected as part of monitoring and assessment activities (i.e. the Far North Caribou Project and Integrated Range Assessments - see Part 2) and the Research Program (see Section 3.2.1). They also worked with landscape attribute data derived from provincial land cover data and disturbance data available for the Hudson Bay Lowlands (HBL) and the developed and undeveloped areas of the boreal forest portion of the Ontario Shield (i.e. DOS and UOS, respectively - Figure 3-13) Ecozone [Project # 50. Please see Appendix 3-1].
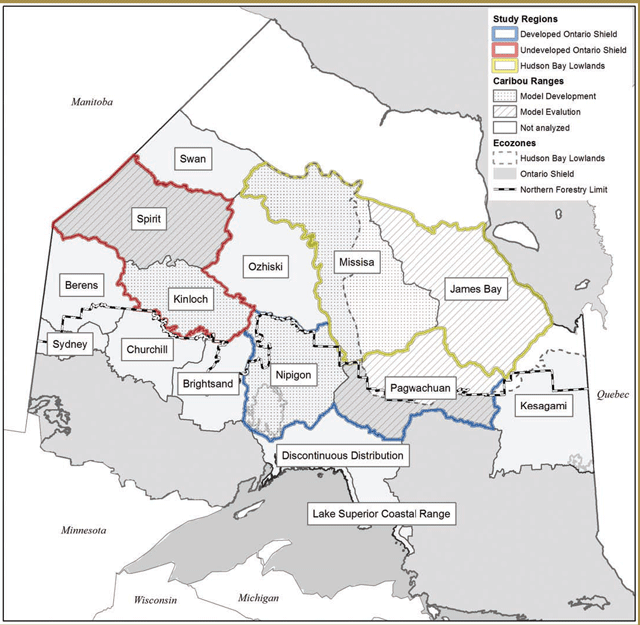
What factors influence broad scale habitat use by caribou?
Researchers used an extensive literature review to develop five alternative hypotheses to explain caribou habitat use across four ranges in the Far North of Ontario (Spirit, Kinloch, Missisa and James Bay) and two more southern ranges (Nipigon and Pagwachuan). These hypotheses were:
- Apparent competition, (ii) Disturbance (both natural and anthropogenic), (iii) Predator avoidance, (iv) Browse availability, and (v) Browse & refuge [Project # 50. Please see Appendix 3-1]. To build upon current research efforts, hypotheses were aligned with those tested under the Research Program (see Section 3.2.1). To test these hypotheses, researchers analyzed areas of high- and low-use by caribou from ranges within HBL, DOS and UOS separately to determine what resources and conditions influence caribou use in these areas. Research focused on:
- Identifying important features associated with caribou use;
- Assessing the likelihood of alternative hypotheses to identify the principal factors that drive caribou habitat selection;
- Determining the scale(s) at which these factors affect caribou habitat selection (i.e. landscape or local);
- Determining whether there are seasonal differences in habitat selection by caribou; and
- Determining the key differences in habitat selection across ranges and RSFs were completed in all of the six above mentioned caribou ranges.
Results from this RSF analysis suggest that forest-dwelling caribou select habitat to reduce predation risk from wolves by avoiding apparent competition with moose and by avoiding disturbed areas. This habitat selection by caribou occurs at broad landscape scales, but patterns of selection differ across ranges and seasons, so multiple RSF models are required for a province-wide assessment of caribou habitat (Figure 3-14). Results also suggest that range boundaries distinguished important patterns in land cover and resource selection. Overall, caribou selected peatlands and coniferous forests while avoiding mixed and deciduous forests, linear features, and naturally disturbed sites.
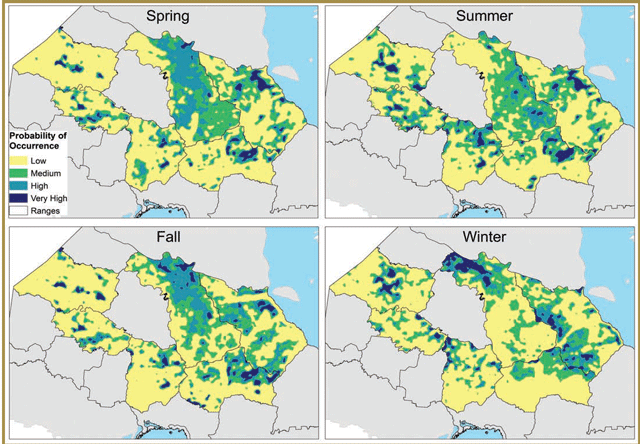
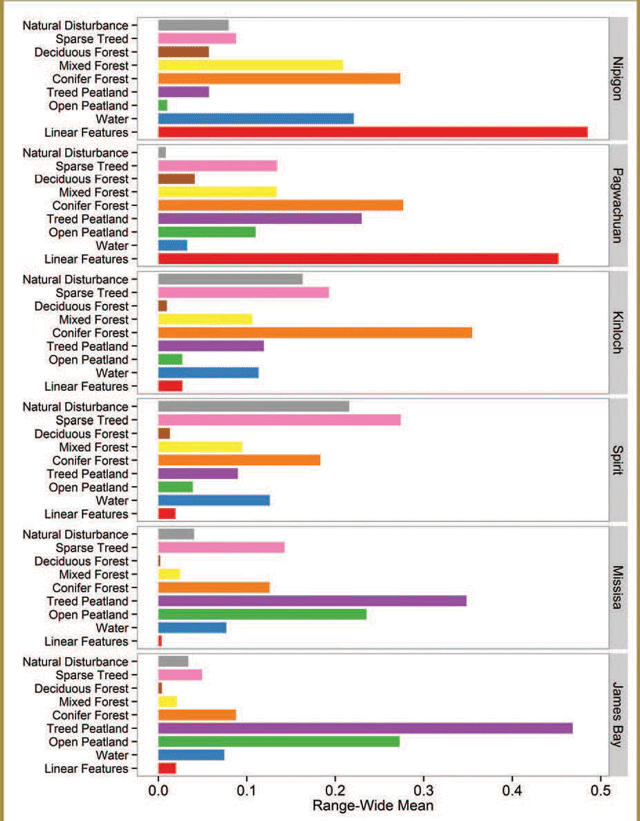
Figure 3-15. Range-wide means of environmental variables across 6 ranges of interest for caribou in northern Ontario. Cover type values are expressed as relative proportion of that cover type within a range, whereas linear features are expressed in terms of density within a range (i.e. km per ha ×10-1).
If you need alternate formats or communication supports to access this, please contact us.
The RSF results have provided insight into the factors that influence caribou use in both the Ontario Shield and the Hudson Bay Lowlands. For example, while habitat use patterns varied between ranges and seasons, predator avoidance was the dominant factor that seemed to influence caribou habitat selection. By documenting the general patterns of selection (e.g. for peatlands and coniferous forests) and avoidance (e.g. of mixed wood and deciduous forests and disturbance and linear features), the RSFs have also allowed researchers to delineate high value caribou habitat within different caribou ranges, within different seasons. As such, the range-specific RSF results can inform planning efforts in these areas. Some potential applications include the following:
- Contributing to the identification of areas that may be sensitive to human activities and development.
- Informing efforts to categorize habitat within caribou ranges (see Section 1.2.3.1 in Part 1).
- Helping identify connected areas that could serve as potential corridors for seasonal caribou migrations.
Additionally, the general RSF approach could be used as a decision support tool to help assess the impacts of human activities on the amount and arrangement of caribou habitat. For example, RSFs can be used to:
- Help assess the potential impacts of different development scenarios on caribou habitat.
- Explore the potential impacts of different climate change scenarios (e.g. a shift to increased moose habitat, particularly in mixed wood forests) on caribou habitat.
- Help model the cumulative effects of different resource sector activities (e.g. forestry and mining activities, utilities and transportation developments) on caribou habitat.
3.3 General overview and synthesis of research findings
3.3.1 Summary of notable research findings and support for research project hypotheses
There are numerous important findings from the extensive caribou research efforts undertaken by MNRF and their partners since the release of the CCP. These include the following:
Ecotype distinction & population structure
- Forest-dwelling and forest-tundra woodland caribou ecotypes have clear behavioural distinctions, although there is geographic overlap in areas used by forest- dwelling and forest-tundra individuals (particularly in winter) (Pond in Berglund et al. 2014).
- Caribou in the Continuous Distribution are distributed in a fairly continuous manner across the Ontario Shield and Hudson Bay Lowlands Ecozones; however, there is evidence for some degree of genetic (Thompson and Wilson in Berglund et al. 2014), behavioural and demographic structure (Shuter & Rodgers 2012) across broader areas.
Population and health measures
- Direct population estimation may be feasible in some situations (e.g. for small, isolated groups – Carr et al. 2012). For situations where it is not feasible or other information is desired (e.g. spatial distribution), there are other metrics that can be used to assess population status and health, each with associated advantages and disadvantages (e.g. occupancy modelling – Poley et al. 2013).
Characterizing habitat
- Caribou habitat selection and time spent in an area (measured at fine and coarse scales) are influenced significantly by both forage availability and predation risk avoidance (Avgar et al. 2013, Avgar et al. submitted, McGreer in prep.).
- Conifer-dominated stands are important to caribou year- round (Avgar et al. 2013, Avgar et al. submitted, McGreer in prep., Poley et al. 2013, Hornseth & Rempel submitted).
- Treed lowlands also have year-round importance for caribou (e.g., Poley et al. 2013, Hornseth & Rempel submitted).
- Caribou avoid habitats that are associated with greater risk of predation or sensory disturbance (e.g. regenerating disturbance, deciduous and mixedwood forests, settled areas, roads) (Poley et al. 2013, Avgar et al. 2013, Hornseth & Rempel submitted).
- There is limited evidence for moderate, fine-scale selection by caribou of regenerating and deciduous stands from November-to April (while regenerating stands are avoided in summer) (Avgar 2013).
Fidelity and space use
- Individual caribou exhibit fidelity to sites used at certain times of the year, as well as both annual and seasonal home Fidelity is strongest to specific sites (Wilson et al. in Berglund et al. 2014) and general areas used during calving (Wilson et al. in Berglund et al. 2014; Sherritt in prep.) and post-calving seasons (Sherritt in prep.).
- In the Far North of Ontario, annual home ranges (i.e. areas used by caribou over the course of a year) were larger in areas with low amounts of preferred habitat (i.e. conifer forest and treed wetlands) and smaller when the amount of suitable caribou habitat was high (Wilson 2013).
- Associated forest and disturbance types had little influence on fidelity to seasonal ranges, with the exception of fidelity to areas used during winter, which was higher when caribou were close to disturbed areas (Sherritt in prep.).
Enhancing regeneration of harvested areas
- Composition of plant communities differed between natural and harvest origin stands and the degree of difference increased with increasing stand age. Differences in forest structure were not as pronounced, but canopy closure was higher in older harvest origin stands (Webster 2013).
- Lichens (particularly ground species) are the dominant component of caribou diets throughout the year (Newmaster et al. 2013; Thompson et al. submitted), but have relatively low nutritional value (Mallon 2014).
- There are several silvicultural techniques that might help promote lichen growth in sites with suitable conditions (e.g. sites with non-organic, sandy soils – McMullin et al. 2011, Kuzyk 2013). These include:
- Re-planting of conifer trees after harvesting (McMullin et al. 2013).
- Careful application of herbicide treatments (i.e. using minimal amounts of types with low lichen impacts) (McMullin et al. 2013, McMullin et al. 2012, McMullin 2011).
- Use of prescribed burning ( vs. mechanical site preparation) (Kuzyk 2013).
- Use of techniques that promote lower tree density and greater canopy openness (e.g. thinning); stand attributes associated with higher lichen abundance (McMullin et al. 2011, Kuzyk 2013).
Re-occupancy or use of harvested areas
- Infrequent examples of caribou use of relatively young harvest origin stands seem more representative of latent use re-occupancy of stands with sufficient regeneration (Preliminary Results).
Energetic balance
- Local environmental conditions (e.g. forage availability, snow depth and temperature), as well as daily movement rates, have a strong influence on caribou energy costs (Mosser et al. 2014).
- Insect harassment appears to be worse in open harvested areas than treed stands (Raponi in prep.). Video evidence for caribou responses to insect harassment suggest that they may reduce time spent feeding when harassment is high, but there does not appear to be a marked increase in energetically costly avoidance behaviours (Preliminary Results).
- Calf and adult condition data was limited, but preliminary evidence for poor calf condition in at least one managed area, combined with relatively low condition indices amongst adults captured in both managed areas, suggest that nutrition may be impacting the health of caribou in some areas (Preliminary Results).
Sensory disturbance
- Caribou may avoid areas with high levels of sensory disturbance (e.g. roads with high traffic volume), while there appears to be less avoidance of roads with lower traffic volumes (Preliminary Results).
- Different road decommissioning strategies have variable levels of success at reducing traffic volume (e.g. seasonal road closures are most effective) (Preliminary Results).
- Along with decommissioning, road reclamation efforts may allow for more effective regeneration of vegetation on roads (Preliminary Results).
Relationships with predators and prey
- Managed areas were found to have higher moose and wolf densities, as well as smaller wolf home range sizes (Anderson 2012, Kittle et submitted, Avgar et al. submitted).
- Moose are the primary year-round prey of wolves (Moffatt 2012, Preliminary Results) and these predators select landscape features that are often associated with higher moose density (i.e. mixedwood, deciduous or disturbed and regenerating forest stands), as well as resource access roads (Anderson 2012, Kittle et submitted).
- The use of resource access roads by wolves enables them to move more rapidly through their territories and improves their hunting success (Moffat 2012, Kittle in prep.).
- Occupancy (Poley et al. 2013), movement behaviour (Avgar et al. 2013, Avgar 2013, Avgar in prep.), habitat selection (Hornseth & Rempel submitted, McGreer in prep.), and activity patterns (Vander Vennen in prep.) of wolves, moose and caribou indicate a high level of overlap between wolves and moose and a high level of spatial and temporal segregation between both of these species and caribou.
- Although caribou are only secondary or tertiary prey items for wolves (Moffatt 2012, Preliminary Results), wolf predation is the dominant cause of adult caribou deaths (Preliminary Results).
Mortality factors
- Caribou in the most intensively managed landscapes have lower survival rates than caribou in the unmanaged landscape and this difference appears to be driven by higher predation rates (Preliminary Results).
- Mortality risk for adult caribou varies seasonally, but patterns of overall and predation-related mortality risk (e.g. low predation rates in summer and relatively high predation rates from late fall to early spring) (Preliminary Results) appear to differ somewhat from seasonal mortality patterns documented in several other jurisdictions (e.g. evidence for peaks in predation and overall mortality rates during summer – Courtois et al. 2007; Rettie and Messier 1998).
- No direct human-caused mortalities of collared caribou were documented in the Research Program study areas (Preliminary Results).
- Most calf deaths in managed and unmanaged landscapes seem to occur within the first two months of life (i.e. from mid-late May to early July) (Preliminary Results), although causes of death have received little direct study and thus, remain.
- Parasite infection levels may be higher amongst caribou in managed unmanaged areas, but research yielded no confirmed evidence of brainworm (P. tenuis) infections (Verocai in prep.).
Population viability analysis (PVA)
- Assessments of probability of population persistence using the simple and spatial caribou PVA models suggest that caribou in one of the managed landscapes may have low probability of persisting over the next 100 years (Preliminary Results).
Resource selection functions (RSFS)
- Selection for habitat is strongest when used and available habitats are characterized at coarse scales (i.e. 10 000 ha) (Hornseth & Rempel submitted).
- While there are some consistent patterns, the selection and avoidance of different habitat types by caribou differs between ranges and seasons (Hornseth & Rempel submitted).
A major objective of many of the research projects described in this section was to assess the degree of supporting evidence (from research results) for alternative hypotheses about how human activities and development affect caribou persistence. While there are several research projects currently underway that will help refine this assessment (see Appendix 3-4 for a detailed list of ongoing research), results obtained to date suggest that there may be support for several of these hypotheses. Table 3-1 summarizes the level and type of support that exists for each hypothesis. Because it appears likely that more than one set of factors are contributing to caribou population declines in managed landscapes (Table 3-1), it can be argued that the Cumulative Effects Hypothesis has the greatest support.
| Hypothesis | General level of support | Relevant evidence | Potential impacts on caribou population health |
|---|---|---|---|
| Energetic balance: Managed forests have lower quality & quantity of caribou food and/or are associated with increased energetic costs of obtaining food. | Moderate |
| Greatest population-level impact may be decreased calf health and possibly, survival rates in managed landscapes |
| Sensory disturbance: Noise and human activity in managed forests influence caribou movement and/or habitat choice. | Moderate |
| Greatest impacts are likely indirect:
|
| Apparent competition: Managed forests attract and support high densities of moose and deer, which results in increased densities of wolves and bears and increased predation rates on caribou. | High | Alternate prey (i.e. moose) and predator (i.e. wolf) densities are higher in managed areas (Kittle et al. submitted, Avgar et al. submitted, Anderson 2012)
| Probable direct impact on adult survival rates
|
| Predator road use: Roads developed in managed forests are used by predators, leading to increased hunting efficiency and higher predation rates on caribou. | Moderate to high |
|
|
| Prey escape: In managed forests, caribou are restricted to fewer and smaller patches of mature conifer forest, which increases search effectiveness of predators. | Low |
|
|
| Cumulative effects: Some combination of two or more of hypotheses contributes to the decline of caribou populations. | Very high |
|
|
In addition to enabling an assessment of the relative degree of support that exists for different hypotheses about factors that underlie caribou population declines, the results of recent research efforts have also provided insights into the biology and ecology of caribou, their predators and a major alternate prey species (i.e. moose). These insights can help inform planning, as well as caribou conservation and/or recovery efforts. Some of the specific implications of research results have been discussed in Section 3.2. However, when all the different research findings are considered at a more general level and in combination with one another, they can also provide broader insights into caribou habitat selection, predator impacts, body condition, use of previously harvested areas, and the potential value of quantitative models (i.e. the PVA and RSF models) as decision support tools.
3.3.2 Caribou habitat
In addition to the project-specific insights into caribou habitat discussed in Section 3.2.4 and in Section 3.2.14, some general insights can also be gained by comparing results obtained from the different habitat selection, RSF and movement studies. Current understandings of caribou habitat have focused on the multi-scale, hierarchical nature of habitat selection (Rettie and Messier 2000). Evidence from several different analyses demonstrates the year-round importance of conifer-dominated forests for caribou at multiple scales (especially the feeding site and stand-levels). Fine scale selection patterns also seem to be influenced by local environmental conditions (e.g. snow depth – Avgar et al. 2013). At the stand and home range scales, caribou also select treed lowlands and avoid or spend less time within disturbed habitats, settled areas and open areas. At all spatial scales, these patterns of habitat selection generally resulted in preference for areas with above average forage availability, but below-average risk of predation, suggesting that both factors may exert a strong influence on caribou habitat selection over a broad range of scales. Results from winter occupancy (Poley et al. 2013) and RSF analyses (Hornseth & Rempel submitted) also suggest that these trends are apparent across both the Ontario Shield and Hudson Bay Lowland Ecozones.
While these general results provide support for current understandings of specific caribou-habitat relationships (e.g. selection for conifer dominated forests, coarse scale avoidance of disturbed areas), some of the habitat research results documented here were not consistent with prior expectations about how caribou respond to different landscape attributes and environmental conditions. For example, the greater abundance and diversity of understory plants available in summer was expected to result in more diversity in caribou diet and selection of foraging habitat. However, analyses of video, GPS collar locations and movement data all suggest that lichen consumption and the use of conifer-dominated stands for foraging are still very important during the summer months. Avoidance of deciduous and regenerating forests by caribou was expected, especially at coarse scales (Rettie and Messier 2000). While such avoidance was observed in several analyses conducted at different scales (e.g. McGreer et al. submitted, Hornseth and Rempel submitted), there was also evidence of a lack of response (i.e. no selection or avoidance) to these cover types at a relatively coarse scale (i.e. monthly movements – Avgar et al. 2013). Additionally, while weaker avoidance or a lack of response to deciduous or regenerating stands by caribou was expected at fi scales (Rettie and Messier 2000), analyses of fi scale movements suggest that during winter, caribou may exhibit weak selection for these stand types (Avgar et al. 2013). Since these habitat types are actively selected by wolves (Kittle et al. submitted, Anderson 2012), such behaviour may place caribou at increased risk of wolf predation.
3.3.3 Predation risk in managed landscapes
Resource management and development can temporarily or permanently convert natural communities into roads or other infrastructure (e.g. mines, work camps) and can alter the amount and distribution of different forest types. These landscape changes have the potential to impact caribou and their habitat. Research results suggest that there are at least two different mechanisms (apparent competition and predator road use) by which the risk of predation-related mortality can increase for caribou that live in managed landscapes.
- Alternate prey and predator densities
Mixedwood and deciduous cover are generally more common in managed boreal landscapes than they are in unmanaged boreal forests that are driven primarily by natural disturbance processes (e.g. wildfire) (Thompson et al. 2003: Table 1). Higher amounts of these forest types, combined with regenerating forests, do appear to support higher densities of alternate prey (e.g. moose) and predators (e.g. wolves) in managed landscapes. Research conducted at fine and coarse spatial scales, indicates that there is a wide degree of separation between the niches of moose and caribou. In contrast, there is a high degree of overlap in niches of wolves and moose (their primary prey), with research indicating that the daily activity levels, habitat selection and spatial distribution of wolves, correspond much more closely with moose than caribou. All of these findings are consistent with the Apparent Competition Hypothesis. Higher predator densities and greater amounts of alternate prey and predator habitat can increase predation risk and potentially, mortality rates for caribou that inhabit managed landscapes.
To mitigate the impacts that changes in the amount and arrangement of different forest types can have on predation risk for caribou, forest management guides in Ontario promote harvesting and silvicultural practices that facilitate the regeneration of conifer-dominated forests to amounts similar to those expected under a natural disturbance regime (MNR 2014, MNR 2010). Conifer regeneration and reduced rate of conversion of conifer-dominated harvested areas to mixedwood and deciduous-dominated stands may result in landscapes with lower alternate prey and predator densities and decrease caribou encounter rates with risky habitat types, all of which have the potential to reduce predation- related impacts on caribou populations.
- Human activities
Research into wolf diets indicates that human garbage associated with communities and work camps (e.g. mining camps, commercial fishing waste disposal sites) can also act as a major food subsidy for wolves (Moffatt 2012, Kittle et al. submitted). Like elevated alternate prey densities, dumpsites may support higher densities of predators in the landscapes they are situated in, which might lead to increased predation-related mortality rates among caribou. Measures that might help mitigate the impacts of dumpsites on caribou populations could include:
- Minimizing the number of dumpsites and/or the quantity of food scraps disposed of in dumpsites; and
- Strategic placement of new dumpsites (e.g. avoidance of high value caribou habitat).
Additionally, habitat selection research suggests that caribou avoid settled areas (e.g. McGreer et al. submitted, Hornseth and Rempel submitted). To help minimize the potential impacts of new settlements (e.g. work camps) on caribou habitat, information on habitat suitability could be used to inform the selection of candidate settlement locations.
- Roads
Roads are another feature of managed landscapes that might have predation-related impacts on caribou.
Preliminary research results suggest that moderate to high traffic volume on some roads may result in fairly strong avoidance of these features, which could lead to impacts like effective habitat loss, habitat fragmentation and reduced connectivity between local populations.
Caribou avoidance of roads with less vehicle traffic was also apparent (e.g. McGreer et al. submitted), however, weaker avoidance of such roads may place caribou at greater risk of predation by wolves. The results of research into wolf habitat selection, suggested that when resource road networks were present, wolves actively selected for them (Anderson 2012, Kittle submitted), likely because these features enabled them to navigate quickly through their territories while targeting suitable moose habitat near forest access roads (Anderson 2012, Kittle submitted). This was further supported by research that indicated that road use and fast travel speeds were the factors with the greatest influence on how efficient wolves were at searching for and capturing their prey (Moffatt 2012). All of these findings are consistent with the Predator Road Use Hypothesis. The positive influence of roads on wolf hunting success and the active selection of roads by wolves, suggests that for caribou or other prey species, the risk of being encountered and predated by wolves increases with proximity to roads. Thus, caribou living in managed landscapes with high road densities might have higher risk of wolf predation and higher mortality rates. The relatively high predation- related mortality rate observed amongst caribou living in a more intensively managed landscape (with relatively high road density) is consistent with this explanation, but this pattern could also be attributed to mechanisms described by other predation-related hypotheses (e.g. Apparent Competition).
Separating the different types of effects that roads might have on caribou (e.g. sensory disturbance impacts vs. predation-related impacts) is important, as potential mitigative measures may be more effective at reducing sensory disturbance-related impacts than they are at reducing predation-related impacts. When feasible, closing or deactivating roads might help reduce the impacts of vehicle traffic on caribou. However, efforts to reduce the functional value of roads as movement corridors that enhance predator hunting success, may also require approaches that re-establish vegetation on road surfaces, in combination with reduced vehicle traffic.
The potential population-level impacts of high traffic volume roads on caribou may be reduced if traffic volumes are reduced (if possible), roads are located in areas with low habitat value (on both sides and at considerable distance from the planned road corridor) and/or if they are located close to a natural subdivision between areas used by different local populations/ demographic units. Efforts to reduce the predation- related impacts that lower traffic volume roads might have on caribou, could include minimizing the extent of their footprint at the landscape level, choosing locations with low habitat value and applying effective road decommissioning and reclamation approaches once roads are no longer needed for resource extraction (e.g. for roads near areas with high suitability for caribou).
In general, applying a road-shed approach to planning and development prior to road building efforts may help minimize the impacts of roads on caribou and reduce costs associated with road decommissioning and rehabilitation. However, additional research would be needed to evaluate the effectiveness of these potential mitigation measures.
- Detection of caribou by predators
Research results obtained to date do not provide much support for the Prey Escape Hypothesis. If development, forest harvesting and other resource extraction activities produce landscapes with less, smaller and more fragmented patches of suitable caribou habitat, there is no strong evidence that predators respond by targeting these areas and the caribou that use them (e.g. due to increased detectability and/or predictability of areas used by caribou). Instead, research results described here are more consistent with a situation where wolves opportunistically kill caribou if they happen to detect or encounter them during targeted searches for their primary prey (i.e. moose). Neither of the managed landscapes studied to date appear to contain wolf packs that actively target and hunt caribou. Instead, the more prominent impact of human activities and developments on the wolf-caribou relationship in these landscapes appears to be the creation of conditions that increase incidental detection of and/or encounters with caribou by wolves.
However, decreases in the number, size and connectivity of areas with suitable caribou habitat may still have negative consequences for the long-term health and persistence of caribou populations. Potential impacts include lower quantities of food (which could lead to poorer caribou condition and lower reproduction and survival rates), lower availability of sites that provide caribou with refuge from areas with high densities of alternate prey and predators, and increased exposure to predation risk when moving between suitable areas. Efforts to produce (at the range level) an amount and arrangement of caribou habitat that is comparable to that expected under a natural disturbance regime (OMNR 2014), in combination with efforts to limit the cumulative amount of disturbance present within a caribou range to levels that are reasonably likely to support a self- sustaining population (i.e. Environment Canada 2008, 2011, MNRF 2014b), may help to reduce some of these impacts.
Higher predation-related and overall mortality rates amongst caribou living in managed landscapes is a prediction shared by all predation-related hypotheses (e.g. Apparent Competition, Predator Road Use and Prey Escape) and there is preliminary evidence that rates of overall and predation-related mortality are higher in landscapes with higher levels of human alteration. Ongoing research into the relationship between predation mortality sites and different landscape attributes will help determine the amount of support for these hypotheses and identify the landscape features associated with greatest risk for caribou.
3.3.4 Reduced condition in managed landscapes (direct and indirect effects)
As discussed above, preliminary results from research conducted to date provides a moderate level of support for both the Energetic Balance and Sensory Disturbance Hypotheses. The Energetic Balance Hypothesis predicts a reduction in net energy gain on caribou populations living in managed forests. The consequent impacts on caribou would likely include reduced body condition, potential decreases in reproductive success (e.g. pregnancy rates and/or live birth rates) and increases in nutrition-related mortality rates amongst adults and/or calves. Direct effects of Sensory Disturbance on caribou may involve behavioural changes (e.g. Section 3.2.8) and elevated stress levels (e.g. Wasser et al. 2011). These responses might lead to the recession and fragmentation of populations (i.e. due to behavioural avoidance of heavily trafficked roads) or reduced body condition due to effective habitat loss and/or increased stress. Again, reduced body condition could also lead to decreases in reproductive success or increases in condition- related mortality rates. Additionally, if caribou health is compromised by nutritional or stress-related factors, they could be more vulnerable to other mortality factors (e.g. predation).
Much of the evidence that supports the Energetic Balance Hypothesis (condition effects: direct) and Sensory Disturbance (possible condition effects: indirect) is preliminary. More comprehensive, finalized research results are needed to determine if there is clear support for preliminary indications that managed landscapes may be associated with reduced caribou condition. If lower body condition is related to a reduction in food availability in managed landscapes, silvicultural treatments that promote the regeneration of ground lichen in harvested areas might be beneficial if applied to suitable sites (i.e. those with non- organic, sandy soils). If lower body condition is influenced by an increase in the energetic costs of acquiring food in managed landscapes, forest management practices that result in landscapes where the amount and arrangement of caribou habitat is comparable to that expected under natural conditions may help reduce the energetic costs of food acquisition and/or finding suitable cover. Additionally, efforts to limit the cumulative amount of disturbance present within a caribou range, to levels that are reasonably likely to support a self-sustaining population (i.e. Environment Canada 2008, 2011, MNRF 2014b) could have a positive influence on food availability and could also help reduce the energetic costs experienced by caribou. Ongoing research into different aspects of caribou energetics should provide some insight into the relative importance of the main factors (i.e. food availability and energetic costs) that are influencing caribou condition in managed landscapes.
3.3.5 Re-occupancy and/or use of previously harvested stands by caribou
Re-occupancy refers to situations where caribou return to areas abandoned after harvesting, following forest regeneration and the development of characteristics associated with suitable habitat (e.g. the growth of conifer- dominated cover and terrestrial lichen) (MNR 2009). This may occur after regenerating stands reach a certain age and in many cases, after appropriate silvicultural treatments have been applied. If caribou are able to successfully re-occupy or use (for cases where prior occupancy of an area is uncertain) previously harvested areas and persist within them, it should be possible to maintain a certain level of forest harvesting in the Continuous and/or Discontinuous Distribution without compromising the long-term viability of affected caribou populations. Despite the importance of the re-occupancy process or caribou use of previously harvested areas, there are several reasons why it is difficult to evaluate with existing data.
First, the term re-occupancy suggests a particular sequence of caribou behaviour in response to harvesting:
Occupancy (prior to harvest)→Abandonment (during and for some time after harvesting) →Re-occupancy/Return to harvested areas (once suitable habitat has regenerated)
Caribou telemetry data are only available for recent years in most areas of Ontario, so while they can help identify caribou use of older and younger harvested stands, they can only be used to identify pre-harvest use and abandonment of recently harvested areas. Since there is a substantial time lag between harvesting or wildfire disturbance and the age when regenerating forests are considered usable by caribou, it is unlikely that telemetry data can be used to document the entire re-occupancy sequence for specific harvested areas.
Consequently, researchers have very limited information on whether older harvested areas were used by caribou before they were harvested, or whether recently harvested areas will be re-occupied by caribou. Due to these challenges, most research efforts will have to focus on recent examples of caribou use of previously harvested areas, without documenting whether this usage constitutes re-occupancy or not.
Second, the results of several studies suggest that caribou select mature, conifer-dominated stands. Current approaches for identifying caribou habitat as part of the forest management planning process do not treat many forest types as suitable until they are ³41 or 61 years old.
The rate at which regenerating forests achieve mature characteristics is affected by harvesting and renewal practices. Modern practices particularly fully mechanized harvesting and processing, followed by the establishment of conifer plantations, did not become common until around 1980, making these kinds of stands < 35 years old. While the equipment used since then has been fairly consistent, forest policy and harvesting techniques have changed considerably over that time period. For example, the maximum size of clear-cuts has changed in response to moose habitat guidelines, and the amount and arrangement of residual trees left standing after harvest has changed in response to the adoption of Natural Disturbance Pattern Emulation Guidelines (MNR 2001). These policies have changed the way regenerating patches are arranged, and may have influenced the effectiveness of some techniques designed to favour conifer regeneration (e.g. aerial application of herbicide). Both factors (i.e. the young age of stands harvested using modern equipment and variation in forest management policy) make it difficult to draw conclusions about the extent to which modern forestry practices are effective at creating future caribou habitat.
A third reason concerns the lack of data on caribou survival, condition and reproductive success in harvested areas. Re- occupancy and/or use of previously harvested areas is an important component of caribou conservation in managed landscapes, but documenting caribou use of harvested areas (and identifying some of the characteristics associated with use) does not address another critical component - the health and long-term viability of caribou that use these areas. Since younger regenerating stands of harvest origin (along with associated road networks) may be associated with higher predation risk, evidence for caribou use of such areas is not necessarily a behaviour that favours caribou persistence (e.g. Dussault et al. 2012). This issue is further complicated by the lack of consensus about how meaningful occupancy by caribou is defined, identified and distinguished from transitory use.
Ongoing research into stand and landscape characteristics associated with caribou re-occupancy of previously harvested stands and caribou mortality rates (Appendix 3-4) may provide insight into the factors that influence re-occupancy and potential consequences that the use of different habitat types might have on caribou survival and population viability.
3.3.6 The PVA & the RSF: Decision support tools for status assessment and scenario analysis
Research can help evaluate support for and expand current understandings of caribou biology and ecology. However, the value of formal research extends beyond documenting evidence for the mere existence of general relationships (e.g. determining whether or not caribou select conifer-dominated forests). The collection of quantitative data can also provide researchers with detailed information regarding the specific form and strength of caribou responses to different landscape attributes or environmental conditions (e.g. a linear or threshold-based relationship between traffic volume and caribou use, or the strength of selection for conifer- dominated forests). It also enables researchers to develop models that can integrate these findings and use them to evaluate the likelihood that caribou will persist over the long- term and on the amount and arrangement of suitable caribou habitat under different development, resource extraction, and climate change scenarios.
For example, the collection of quantitative data on factors that influenced caribou habitat selection and movements allowed researchers to develop a detailed spatial simulation model that makes predictions about long-term changes in caribou population sizes (i.e. the spatially-explicit PVA model - Section 3.2.13). This model can serve as a powerful decision support tool for assessing the cumulative effects of existing disturbance and developments on caribou population viability and for comparing the potential impacts of different planning scenarios on caribou persistence (see Section 3.2.13). The collection of detailed, quantitative data on different factors that influence caribou habitat use throughout multiple ranges (including several in the Far North of Ontario) enabled researchers to develop and apply RSFs to spatially delineate suitable caribou habitat in different ranges and during different seasons. Spatial maps of suitable caribou habitat can be used to identify areas that might be sensitive to development and resource extraction activities, as well as connected areas that could serve as seasonal migration corridors.
In addition to the contribution the PVA and RSF models can make to informing assessments of the current state of caribou populations and habitat, these models could also be applied to simulated landscapes that represent different planning, development and/or climate change scenarios. The results could help evaluate and compare the potential impacts of these scenarios on caribou populations and habitat. The process of developing and evaluating alternative representations of future landscapes can enable researchers to incorporate alternative perspectives and explore the potential cumulative impacts of different developments and/or landscape changes on caribou and their habitat. As such, they can be used to improve understandings of key uncertainties. The PVA modelling framework described in Section 3.2.13 is very flexible in that it provides opportunities to explore the impacts of new scenarios on the probability of caribou persistence as they arise or to incorporate new information as it becomes available. Similarly, the RSF models described in Section 3.2.14 can also be applied to delineate the amount and arrangement of suitable caribou habitat in different ranges, under different scenarios. PVA and /or RSF results obtained under alternative scenarios can be compared and used to rank alternative planning and development options in relation to the level of their potential impacts on caribou persistence and/or habitat. New decision support tools generated from research and monitoring work can be useful in caribou conservation and recovery planning efforts (MNR 2009; Action 7.4).
For example, the spatial PVA model has some key features that allow users to conduct comprehensive and detailed assessments of the potential impacts new developments or disturbances might have on the long-term viability of different caribou populations. It can be used to simultaneously model the effects of different disturbance and activity types on caribou movement, habitat selection and mortality rates. The cumulative effects of disturbance can be integrated together, over time, allowing users to assess their combined impacts on the probability that caribou populations will persist over the long-term. The spatial PVA model also allows users to differentiate between different disturbance types – enabling them to directly incorporate differences in the documented effects they might have on caribou movement, habitat selection and mortality risk.
Finally, the PVA can be used as a tool for directly modelling the impacts of different non-spatial factors (e.g. changes in moose and/or wolf harvest rates) on caribou population viability. This feature enables the direct evaluation of the potential consequences that implementing different combinations of spatial and non-spatial planning options (e.g. different forest harvest rotation times and cutblock dimensions, different road development scenarios, different levels of moose hunting) might have on caribou population persistence.
3.4 Knowledge gaps and future research
Finally, while completed and ongoing research efforts (see detailed list in Appendix 3-4) are reducing some of the key uncertainties related to the impacts of anthropogenic disturbance on caribou populations, and the effectiveness of different measures that might help mitigate those impacts, there are some major knowledge gaps that will still need to be addressed once all planned research efforts are finished. However, conducting research to address these gaps may be quite challenging (e.g. due to required resources and animal care concerns) and would require different data types, sampling equipment and sampling strategies than those used to date.
Major gaps include:
- Direct information on the primary causes of caribou calf mortality;
- A comprehensive assessment of the role of black bears as a cause of caribou calf mortalities and an assessment of how bear density, habitat selection and feeding behaviour are impacted by anthropogenic disturbances in the Continuous and/or Discontinuous Distribution;
- Detailed information on the causes of adult caribou deaths that are not predation-related; and
- Long-term study of caribou abandonment and re-occupancy of previously harvested
In addition to these knowledge gaps, logistical and funding constraints meant that several of the Research Program projects described here, were restricted to only one or two study areas. In these cases, study areas were selected to maximize managed vs. unmanaged contrasts or to obtain a detailed picture of the ecology of a highly managed landscape. Even for those projects that were able to use data from three different areas, the lack of multiple example landscapes or replicates for each management category creates uncertainty regarding the representativeness of research findings for managed and unmanaged landscapes throughout the Continuous Distribution of caribou in Ontario. To improve confidence in the extent to which research results are representative of caribou biology and ecology throughout the Continuous Distribution, studies would need to be expanded to include data from all three study areas. Ideally, data from other managed and unmanaged study areas would also need to be collected and analyzed.
Finally, a synthesis of research results described in this report suggests that the level of support for the six alternative hypotheses initially identified by the Research Program is variable (Table 3-1). A potential framework for future caribou research could involve further testing of the hypotheses associated with a considerable degree of supporting evidence (e.g. Apparent Competition,
Predator Road Use, Sensory Disturbance, and Energetic Balance). Several different measures have been suggested as possible options for mitigating some of the direct and indirect impacts that human activities might have on caribou populations (see Section 3.3). However, in most cases, the effectiveness of many of these measures (in terms of direct impacts on caribou behaviour and/or vital rates) has not been tested. Future research efforts could focus on evaluating how effective different management actions are at reducing potential impacts of key mechanisms that influence the viability of caribou populations. The results of this type of approach can provide greater certainty regarding the impacts of different caribou-oriented conservation and recovery planning actions (e.g. road decommissioning, different harvesting techniques) on caribou persistence. Research results could also continue to improve understandings of the general factors that influence caribou persistence.
References
Anderson, M.L. 2012. Wolf responses to spatial variation in moose density in northern Ontario. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada.
Andrén, H., J.D.C. Linnell, O. Liberg, R. Andersen, A. Danell, J. Karlsson, J. Odden, P.F. Moa, P. Ahlqvist, T. Kvam, R. Franzén, P. Segerström. 2006. Survival rates and causes of mortality in Eurasian lynx (Lynx lynx) in multi- use landscapes. Biological Conservation. 131: 23-32.
Avgar, T. 2013. From Diffusion to Cognition: Analytical, Statistical and Mechanistic Approaches to the Study of Animal Movement. Ph.D. Thesis. University of Guelph, Guelph, ON, Canada.
Avgar, T., R. Deardon and J.M. Fryxell. 2013. An empirically parameterized individual based model of animal movement, perception, and memory. Ecological Modelling. 251: 158-172.
Avgar, T., A. Mosser, G.S. Brown & J.M. Fryxell. 2013. Environmental and individual drivers of animal movement patterns across a wide geographical gradient. Journal of Animal Ecology. 82(1): 96-106.
Avgar, T., J. Baker, G.S. Brown, J. Hagens, E. Iwachewski, A. Kittle, E. Mallon, M. McGreer, A. Mosser, S.G. Newmaster, B. Patterson, D. Reid, A. Rodgers, J. Shuter, G. Street, I. Thompson, M. Turetsky, P. Wiebe, and J.M. Fryxell. (submitted manuscript). Space-use behaviour of woodland caribou based on a cognitive movement model.
Bergerud, A.T. 1972. Food habits of Newfoundland caribou. Journal of Wildlife Management. 36: 913-923. Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp.
Brown, K.M. 1999. Planning the next round of caribou habitat research: Results of the workshop held in Thunder Bay, Ontario 29 and 30 April, 1999. Sponsored by Forest Ecosystem Science Co-operative Inc., Thunder Bay, Ontario. 44 pp. 127.
Brown, G.S., L. Landriault. D.J.H. Sleep and F.F. Mallory. 2007. Comment arising from a paper by Wittmer et al.: hypothesis testing for top-down and bottom-up effects in woodland caribou population dynamics. Oecologia. DOI 10.1007/s00442-007-0855-3.
Canadian Broadcasting Corporation (CBC). 2007 (Apr. 30). Poaching investigated in threatened caribou herds.
Carr, N.L., A.R. Rodgers, S.R. Kingston, P.N. Hettinga, L.M. Thompson, J.L. Renton and P.J. Wilson, 2012. Comparative woodland caribou population surveys in Slate Islands Provincial Park, Ontario. Rangifer. 32(2), 205-218.
Caughley, G. and A. Sinclair. 1994. Wildlife Ecology and Management. Blackwell Science: Cambridge, MA.
Courtois, R., J.P. Ouellet, L. Breton, A. Gingras, and C. Dussault. 2007. Effects of forest disturbance on density, space use, and mortality of woodland caribou. Ecoscience. 14(4): 491-498.
Darby, W. R., & Duquette, L. S. (1986). Woodland caribou and forestry in Northern Ontario, Canada. Rangifer. 6(2):87-93.
Downes, C.M., J. B. Theberge and S. M. Smith. 1986. The influence of insects on the distribution, microhabitat choice and behavior of the Burwash caribou herd. Canadian Journal of Zoology. 64: 622-629.
Dussault, C., V. Pinard, J.P. Ouellet, R. Courtois and D. Fortin. 2012. Avoidance of roads and selection for recent cutovers by threatened caribou: fitness-rewarding or maladaptive behaviour? Proceedings of the Royal Society B. rspb20121700.
Environment Canada. 2008. Scientific review for the identification of critical habitat for woodland caribou (Rangifer tarandus caribou), Boreal Population, in Canada. August 2008. Ottawa: Environment Canada. 72 pp. + appendices.
Environment Canada. 2011. Scientific Assessment to inform the identification of critical habitat for woodland Caribou (Rangifer tarandus caribou), Boreal population, in Canada; update. Ottawa, Ontario, Canada. 102 pp. + appendices. Environment Canada, & Agriculture and Agri-Food Canada.
Faille, G., C. Dussault, J.P. Ouellet, D. Fortin, R. Courtois, M.-H. St-Laurent and C. Dussault. 2010. Range fidelity: the missing link between caribou decline and habitat alteration?. Biological Conservation. 143(11): 2840-2850.
Ferguson, S.H., and P.C. Elkie. 2004. Seasonal movement patterns of woodland caribou (Rangifer tarandus caribou). Journal of Zoology 262(2): 125 – 134.
Gaillard, J.-M., M. Festa-Bianchet and N.G. Yoccoz. 1998. Population dynamics of large herbivores: variable recruitment with constant adult survival. Trends in Ecology & Evolution 13(2): 58-63.
Gustine, D.D., K.L. Parker, R.J. Lay, M.P. Gillingham and D.C. Heard. 2006. Calf survival of woodland caribou in a multi-predator ecosystem. Wildlife Monographs. 165:1-32.
Hornseth, M.L. and R. S. Rempel. (submitted manuscript). Identifying regional differences in habitat selection of Boreal Woodland Caribou (Rangifer tarandus caribou): A range-based approach.
Jacques, C.N., T.R. Van Deelen, W.H. Hall Jr., K.J. Martin and K.C. Vercauteren. 2011. Evaluating How Hunters See and React to Telemetry Collars on White-Tailed Deer. The Journal of Wildlife Management. 75(1): 221- 231.
Kittle, A.M., B.R. Patterson, M.L. Anderson, S. Moffatt A.R. Rodgers, J.L. Shuter, D.E.B. Reid, J.A. Baker, G.S. Brown, I.D. Thompson, G.M. Street, T. Avgar, L. Vander Vennen, A. Mosser, J. Hagens, E Iwachewski and J.M. Fryxell. (submitted manuscript). Wolves adapt territory size, not pack size to local habitat quality.
Kittle, A.M. (in prep.). The impact of prey, habitat and anthropogenic disturbance on space use by social carnivores. PhD thesis, University of Guelph.
Klütsch C.F.C., M. Manseau and P.J. Wilson. 2012. Phylogeographical Analysis of mtDNA Data Indicates Postglacial Expansion from Multiple Glacial Refugia in Woodland Caribou (Rangifer tarandus caribou). PLoS one 7(12): e52661. doi:10.1371/journal.pone.0052661
Kuzyk, R.E. 2013. Terrestrial lichen abundance in relation to stand structure and silvicultural history. M.Sc. Thesis. Lakehead University, Thunder Bay, ON, Canada Liberg, O., G. Chapron, P. Wabakken, H.C. Pedersen, N.T. Hobbs and H. Sand. 2012. Shoot, shovel and shut up: cryptic poaching slows restoration of a large carnivore in Europe. Proceedings of the Royal Society B: Biological Sciences. 279(1730): 910-915.
Lowes, K. 2013. Body condition of woodland caribou (Rangifer tarandus caribou) in managed and unmanaged Ontario forests. HBEM. Thesis. Lakehead University, Thunder Bay, ON, Canada.
Martin-DeMoor, J. V.J. Lieffers and S.E. Macdonald. 2010. Natural regeneration of white spruce in aspen-dominated boreal mixedwoods following harvesting. Canadian Journal of Forest Research 40: 585-594.
Mallon, E. 2014. Effects of Disturbance and Landscape Position on Vegetation Structure and Productivity in Ontario Boreal Forests: Implications for woodland caribou (Rangifer tarandus caribou) Forage. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada.
Moffatt, S. 2012. Time to event modelling: wolf search efficiency in northern Ontario. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada.
Mosser, A., T. Avgar, G.S. Brown, C.S. Walker and J.M. Fryxell. 2014. Towards an energetic landscape: broad- scale accelerometry in woodland caribou. Journal of Animal Ecology. (Early View currently available online – DOI 10.1111/1365-2656.12187).
McGreer, M. (in prep.). Influence of spatial variation in food availability and predation risk on habitat selection by woodland caribou in Ontario. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada.
McGreer, M., T. Avgar, J. Baker, G.S. Brown, J. Hagens, E. Iwachewski, A. Kittle, E. Mallon, A. Mosser, S.G. Newmaster, B. Patterson, D. Reid, A. Rodgers, J. Shuter, G. Street, I.D. Thompson, M. Turetsky, L.M. Vander Vennen, P. Wiebe and J.M. Fryxell. (submitted manuscript). Influence of spatial variation in forage availability and predation risk on habitat selection by woodland caribou (Rangifer tarandus caribou) in Ontario.
McLellan, B.N., F.W. Hovey, R.D. Mace, J.G. Woods, D.W. Carney, M.L. Gibeau and W.F. Kasworm. 1999. Rates and causes of grizzly bear mortality in the interior mountains of British Columbia, Alberta, Montana, Washington, and Idaho. The Journal of Wildlife Management. 63(3): 911-920.
McMullin, R.T. 2011. Relationships between Silviculture, Lichen Diversity and Woodland Caribou (Rangifer tarandus caribou) in Northern Ontario. Ph.D. Thesis. University of Guelph, Guelph, ON, Canada.
McMullin, R.T., F.W. Bell, S.G. Newmaster. 2012. The effects of triclopyr and glyphosate on lichens. Forest Ecology and Management. 264(15): 90-97.
McMullin, R.T., I.D. Thompson, S.G. Newmaster. 2013. Lichen conservation in heavily managed boreal forests. Conservation Biology. 27(5): 1020-30.
McMullin, R.T., I.D. Thompson, S.G. Newmaster, B.W. Lacey. 2011. Estimating the biomass of woodland caribou forage lichens. Canadian Journal of Forest Research. 41: 1961-9.
Ministry of Natural Resources (MNR). 2001. Forest Management Guide for Natural Disturbance Pattern Emulation – Technical Series, Version 3.1. Queen’s Printer for Ontario, Toronto. 40 pp.
MNR. 2009. Ontario’s Woodland Caribou Conservation Plan. Queen’s Printer for Ontario, Toronto Ontario, Canada. 24 pp.
MNR. 2010. Forest Management Guide for Conserving Biodiversity at the Stand and Site Scales. Queen’s Printer for Ontario, Toronto, Ontario, Canada. 211 pp.
MNR. 2014. Forest Management Guide for Boreal Landscapes. Queen’s Printer for Ontario, Toronto, Ontario, Canada. 104 pp.
Ministry of Natural Resources and Forestry (MNRF). 2014a. Delineation of Woodland Caribou Ranges in Ontario. MNRF, Species at Risk Branch, Thunder Bay, Ontario.
MNRF. 2014b. Range Management Policy in Support of Woodland Caribou Conservation and Recovery. Species at Risk Branch, Thunder Bay, Ontario. 11 pp.
Newmaster, S.G., R.A.D. Steeves, A.R. Rodgers, A.J. Fazekas, J.R. Maloles, R.T. McMullin and J.M. Fryxell. 2013. Examination of two new technologies to assess the diet of woodland caribou: video recorders attached to collars and DNA barcoding. Canadian Journal of Forest Research. 43:897-900.
Newton E.J. 2012. Factors affecting changes in the distribution and abundance of migratory caribou (Rangifer tarandus) in the Hudson Bay Lowlands. MSc thesis, Trent University, Peterborough, Ontario, Canada.
Newton, E.J., B.A. Pond, G.S. Brown, K.F. Abraham, J.A. Schaefer. 2014. Remote sensing reveals long-term effects of caribou on tundra vegetation. Polar Biology. 37(5): 715-725.
Newton, E.J. K.F. Abraham, J.A. Schaefer, B. A. Pond, G.S. Brown, and J.E. Thompson. In revision. Broad- scale changes in the distribution of migratory caribou (Rangifer tarandus) of southern Hudson Bay: effects of forage and human disturbance. Arctic.
Ontario Ministry of Natural Resources (OMNR). 2014. Forest Management Guide for Boreal Landscapes. Queen’s Printer for Ontario, Toronto. 101 pp.
Poley, L.G., B.A. Pond, J.A. Schaefer, G.S. Brown, J.C. Ray and D.S. Johnson, 2013. Occupancy patterns of large mammals in the Far North of Ontario under imperfect detection and spatial autocorrelation. Journal of Biogeography. 1-11.
Pond, B.A. Ecotype differentiation. pp. 43-49. in Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp.
Pinard, V., C. Dussault, J-P Ouellet, D. Fortin, R. Courtois. 2012. Calving Rate, Calf Survival Rate, and Habitat Selection of Forest-Dwelling Caribou in a Highly Managed Landscape. The Journal of Wildlife Management. 76(1):189-199.
Ontario Ministry of Natural Resources (OMNR). 2013. Provincial Satellite Derived Disturbance Mapping Data Specifications. Inventory, Monitoring and Assessment Section Technical Report (Unpublished) 20p.
Qi, M. and J.B. Scarratt. 1998. Effect of harvesting method on seed bank dynamics in a boreal mixedwood forest in northwestern Ontario. Canadian Journal of Botany 76: 872-883.
Raponi, M. (in prep.). Role of biting flies as a source of harassment for woodland caribou in Ontario. M.Sc. Thesis. Trent University, Peterborough, ON, Canada.
Rettie, W. and F. Messier. 1998. Dynamics of woodland caribou populations at the southern limit of their range in Saskatchewan. Canadian Journal of Zoology. 76: 251-259.
Rettie, W. J., and F. Messier. 2000. Hierarchical habitat selection by woodland caribou: its relationship to limiting factors. Ecography. 23(4): 466-478.
Rodgers, A.R., B.A. Allison, K.D. Wade and E.P. Iwachewski. Forest-Dwelling Woodland Caribou in Ontario: Research Workshop Report. Information Paper CNFER IP-001. Ontario Ministry of Natural Resources, Centre for Northern Forest Ecosystem Research, Thunder Bay, Ontario, Canada. 27pp.
Rodgers, A.R., N.E. Berglund, K.D. Wade, B.A. Allison and E.P. Iwachewski. 2008. Forest-Dwelling Woodland Caribou in Ontario: Experimental Design Workshop Report. CNFER Information Paper IP-002. Ontario Ministry of Natural Resources, Centre for Northern Forest Ecosystem Research, Thunder Bay, Ontario, Canada. 19pp.
Rodgers, A.R., N.E. Berglund, J.E. Hagens, K.D. Wade, B.A. Allison and E.P. Iwachewski. 2009. Forest-Dwelling Woodland Caribou in Ontario: Candidate Study Areas Report. CNFER Information Paper IP-004. Ontario Ministry of Natural Resources, Centre for Northern Forest Ecosystem Research, Thunder Bay, Ontario, Canada. 48pp.
Wasser. S.K., J.L. Keim, M.L. Taper and S.R. Lele. 2011 The influences of wolf predation, habitat loss, and human activity on caribou and moose in the Alberta oil sands. Frontiers in Ecology and the Environment. 9(10): 546–551, doi:10.1890/100071.
Rudolph. T.D., P. Drapeau, M-H. St-Laurent, L. Imbeau. 2012. Status of Woodland Caribou (Rangifer tarandus caribou) in the James Bay Region of Northern Quebec. Scientific report presented to the Ministère des Ressources naturelles et de la Faune du Québec and the Grand Council of the Crees (Eeyou Istchee). Montreal, QC. 72 pp.
Sherritt, A. (in prep.). Rapid assessment of woodland caribou habitat: the influence of habitat on site fidelity. M.Sc. Thesis. Trent University, Peterborough, ON, Canada.
Thompson, I.D., P.A. Wiebe, E. Mallon, A.R. Rodgers, J.M. Fryxell, J.A. Baker. (submitted manuscript). Factors influencing the seasonal diet selection by woodland caribou in boreal forests in Ontario.
Thompson, L. (in prep.). Ph.D. Thesis. Trent University, Peterborough, ON, Canada.
Thompson, L., and P.J. Wilson. (in prep). Genetic Structure in Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Ministry of Natural Resources & Forestry.
Thompson, L., and P.J. Wilson. Genetic structure. pp. 83-88 in Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp.
Toupin, B., J. Huot and M. Manseau. 1996. Effect of Insect Harassment on the Behaviour of the Rivière George Caribou. Arctic. 49(4):375-382.
Shuter, J.L., & Rodgers, A. R. 2012. Delineating demographic units of woodland caribou (Rangifer tarandus caribou) in Ontario: cautions and insights. Rangifer. 32(2), 159-181.
Thompson, I. D., J. A. Baker and M. Ter-Mikaelian. 2003. A review of the long-term effects of post-harvest silviculture on vertebrate wildlife, and predictive models, with an emphasis on boreal forests in Ontario, Canada. Forest Ecology and Management. 177(1): 441-469.
Thompson, I.D., M. Bakhtiari, A.R. Rodgers, J.A. Baker, J.M. Fryxell, E. Iwachewski. 2012. Application of a high- resolution animal-borne remote video camera with global positioning for wildlife study: Observations on the secret lives of woodland caribou. Wildlife Society Bulletin. 36(2):365-70.
Thompson, I.D., P.A. Wiebe, E. Mallon, A.R. Rodgers, J.M. Fryxell, J.A. Baker. (submitted manuscript). Factors influencing the seasonal diet selection by woodland caribou in boreal forests in Ontario.
Vander Vennen, L. (in prep.). Diel variability of wolf predation on ungulate prey. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada.
Verocai, G. (in prep.). Ph.D. Thesis. University of Calgary, Calgary, AB, Canada.
Wasser, S.K., J.L. Keim, M.L. Taper, R.L. Subhash. 2011. The influences of wolf predation, habitat loss, and human activity on caribou and moose in the Alberta oil sands. Frontiers in Ecology and the Environment. doi:10.1890/100071.
Whittington, J., M. Hebblewhite, N.J. DeCesare, L. Neufeld, M. Bradley, J. Wilmshurst, and M. Musiani. 2011. Caribou encounters with wolves increase near roads and trails: a time-to-event approach. Journal of Applied Ecology 48(6): 1535-1542.
Wilson, K.S. 2013. Temporal and spatial variation in home range size for two woodland caribou ecotypes in Ontario. M.Sc. Thesis. Trent University, Peterborough, ON, Canada.
Wilson, K., G.S. Brown and B.A. Pond. Movement and activity analysis home range and season delineation). pp. 50-54 in Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp.
Wilson, P.J. and L. Thompson. 2008. A preliminary genetic assessment of forest-dwelling and forest-tundra populations of woodland caribou in Ontario, Canada. Unpublished report to the Ontario Ministry of Natural Resources. May 2008. 10 pp.
Wittmer, H., A. Sinclair and B. McLellan. 2005. The role of predation in the decline and extirpation of woodland caribou. Oecologia. 144:257-267.
Webster, N.T.W. 2013. Forest management does not emulate natural disturbance with respect to plant diversity and forest community composition. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada.
Appendix 3-1: General summary of recent research projects that focus on woodland caribou in Ontario
| Project # | Section in this report | Research initiative | Specific research focus | Location | Hypothesis tested | CCP Action item | Project leads (People), Organization affiliation |
|---|---|---|---|---|---|---|---|
| 1 | 3.2.2 | Monitoring and Assessment | Caribou ecotype distinction | Far North of Ontario | A clear behavioural and spatial distinction can be made between the forest-tundra and forest- dwelling ecotypes of woodland caribou | 1.1, 2.2, 7.4 | Avgar, T., Univ. of Guelph Fryxell, J.M., Univ. of Guelph Brown, G.S., MNRF |
| 2 | 3.2.2 | Monitoring and Assessment | Caribou ecotype distinction | Far North of Ontario | A clear behavioural and spatial distinction can be made between the forest-tundra and forest- dwelling ecotypes of woodland caribou | 1.1, 2.2, 7.4 | Pond, B., MNRF Wilson, K., Trent University Schaefer, J., Trent University Brown, G.S., MNRF |
| 3 | 3.2.2 | Range Delineation (historic telemetry data – collected prior to 2009) | Caribou population delineation | Southern portion of the Continuous Distribution of caribou, within the area of current commercial harvesting | The distribution of caribou in Ontario is continuous, but spatially structured and comprised of multiple local populations with overlapping ranges | 1.1, 2.2, 7.4 | Shuter, J.L., MNRF Rodgers, A.R., MNRF |
| 4 | 3.2.2 | Monitoring and Assessment | Caribou population delineation | The Continuous Distribution of caribou (excluding Lake Superior Coast Range) | The distribution of caribou in Ontario is continuous, but spatially structured and comprised of multiple local populations with overlapping ranges A clear behavioural and spatial distinction can be made between the forest-tundra and forest- dwelling ecotypes of woodland caribou | 1.1, 2.2, 7.4 | Brown, G.S., MNRF |
| 8 | 3.2.3 | Collaborative Woodland Caribou Research Program | Testing predictions that different hypothesis make regarding individual-level condition and population level vital rates | Pickle Lake, Nakina and Cochrane study areas | All major hypotheses assessed as part of the Collaborative Woodland Caribou Research Program (see Section 3.2.1) | 1.1, 1.4 | Rodgers, A.R., MNRF Fryxell, J.M., Univ. of Guelph Brown, G.S., MNRF Thompson, I.D., Canadian Forest Service (CFS) |
| 9 | 3.2.4 | Collaborative Woodland Caribou Research Program | Determining which habitats caribou select when feeding in managed and unmanaged forests | Pickle Lake, Nakina and Cochrane study areas | Habitat selection is driven by forage quantity/quality or Habitat selection is driven by predation risk avoidance | 1.2, 4.1, 4.2 | Thompson, I.D., CFS |
| 10 | 3.2.4, 3.2.6 | Collaborative Woodland Caribou Research Program | Determining which cover types and aspects of forest stand structure are associated with summer caribou use | Pickle Lake and Nakina study areas | Habitat selection is driven by forage quantity/quality Habitat selection is driven by predation risk avoidance | 1.2, 4.1, 4.2 | Reid, D., MNRF Buda, N., MNRF |
| 11 | 3.2.4, 3.2.13, 3.2.14 | Collaborative Woodland Caribou Research Program | Determining how caribou movement is affected by different landscape attributes and environmental conditions | Far North of Ontario and southern portion of the Continuous Distribution | Habitat selection is driven by forage quantity/quality Habitat selection is driven by predation risk avoidance | 1.2, 4.1, 4.2, 5.2, 5.4, 5.5 | Avgar, T., Univ. of Guelph McGreer, M., Univ. of Guelph Mosser, A., Univ. of Guelph Brown, G.S., MNRF Fryxell, J.M., Univ. of Guelph |
| 12 | 3.2.4 | Monitoring and Assessment | Determining whether different forest and disturbance types affect fidelity to areas used by caribou in different seasons | Cochrane and 2 additional study locations in Southern Manitoba | Traditional use of seasonal areas is driven by individual familiarity Traditional use of seasonal areas is influenced by individual familiarity but is also responsive to variation in habitat suitability | 1.2, 4.1, 4.2 1.2, 4.1, 4.2 | Sherritt, A., Trent University Schaefer, J., MNRF Pond, B. , Trent University Brown, G.S., MNRF |
| 13 | 3.2.4 | Monitoring and Assessment | Determining whether the level of fidelity individuals show towards calving sites differs between forest- dwelling and forest- tundra woodland caribou | Far North of Ontario | Wilson, K., Trent University Brown, G.S., MNRF Pond, B., MNRF | ||
| 14 | 3.2.4 | Monitoring and Assessment | Determining whether home range sizes are affected by different landscape attributes and environmental conditions | Far North of Ontario | Home range characteristics of the two woodland caribou ecotypes differ and these differences are influenced by variation in resources and environmental conditions | 1.2, 4.1, 4.2 | Wilson, K., Trent University Schaefer, J., MNRF Pond, B., Trent University Brown, G.S., MNRF |
| 15 | 3.2.5 | Collaborative Woodland Caribou Research Program | Determining if plant communities differ between natural and harvest-origin stands of different ages | Pickle Lake and Nakina study areas | Forest harvesting emulates natural disturbances, in terms of the composition and structure of the plant community that develops after disturbance | 1.2 | Webster, N., Univ. of Guelph Newmaster, S.G., Univ. of Guelph Reid, D., MNRF |
| 16 | 3.2.5 | Collaborative Woodland Caribou Research Program | Determining how different forest characteristics affect ground lichen abundance | Pickle Lake and Nakina study areas | Multiple hypotheses about the conditions, resources and processes that influence lichen growth | 1.2 | Kuzyk, R., Lakehead Univ. Reid, D., MNRF |
| 17 | 3.2.5 | Collaborative Woodland Caribou Research Program | Determining how different forest characteristics affect ground and tree lichen diversity | Several managed forests in northwestern and northeastern Ontario | Multiple hypotheses about the conditions, resources and processes that influence lichen growth | 1.2 | McMullin, T., Univ. of Guelph Thompson, I.D., CFS Newmaster, S.G., Univ. of Guelph |
| 18 | 3.2.5 | Collaborative Woodland Caribou Research Program | Determining the impacts of different silvicultural treatments on ground lichen communities | Nakina study area | Multiple hypotheses about the conditions, resources and processes that influence lichen growth | 1.2 | Kuzyk, R., Lakehead Univ. Reid, D., MNRF |
| 19 | 3.2.5 | Collaborative Woodland Caribou Research Program | Determining the impacts of different herbicide treatments on ground and tree lichen communities | Cochrane study area | Multiple hypotheses about the conditions, resources and processes that influence lichen growth | 1.2 | McMullin, T., Univ. of Guelph Bell, F.W., MNRF Newmaster, S.G., Univ. of Guelph Thompson, I.D., CFS |
| 20 | 3.2.5 | Collaborative Woodland Caribou Research Program | Determining the impacts of different silvicultural treatments on ground and tree lichen communities | Several sites in northwestern Ontario and several in the Cochrane study area | Multiple hypotheses about the conditions, resources and processes that influence lichen growth | 1.2 | McMullin, T., Univ. of Guelph Thompson, I.D., CFS Newmaster, S.G., Univ. of Guelph |
| 21 | 3.2.6 | Collaborative Woodland Caribou Research Program | Identifying caribou use of previously harvested areas & determining what factors influence re-occupancy | Nakina study area | Multiple hypotheses about stand age, type and structural characteristics that might influence caribou re-occupancy | 1.2 | Reid, D.,MNRF Buda, N., MNRF |
| 22 | 3.2.7 | Collaborative Woodland Caribou Research Program | Characterizing caribou diets – measuring the composition and relative abundance of different forage species | Pickle Lake, Nakina and Cochrane study areas | Energetic Balance Hypothesis (Section 3.2.1): (focus: energy intake component) | 1.1, 4.1 | Thompson, I.D., CFS Newmaster, S.G., Univ. of Guelph Rodgers, A.R., MNRF |
| 23 | 3.2.7 | Collaborative Woodland Caribou Research Program | Characterizing nutritional value of caribou diets | Pickle Lake, Nakina and Cochrane study areas | Energetic Balance Hypothesis: (focus: energy intake component) | 1.1, 4.1 | Mallon, E., Univ. of Guelph Turetsky, M.R., Univ. of Guelph Thompson, I.D., CFS |
| 24 | 3.2.7 | Collaborative Woodland Caribou Research Program | Determining the influence of different behaviours and environmental conditions on caribou energy expenditures | Pickle Lake, Nakina and Cochrane study areas, Toronto Zoo | Energetic Balance Hypothesis: (focus: energy expenditure component) | 1.1, 4.1 | Mosser, A., Univ. of Guelph Fryxell, J.M., Univ. of Guelph |
| 25 | 3.2.7 | Collaborative Woodland Caribou Research Program | Developing models for snow accumulation and melting in different forest types and assessing snow impacts on caribou energy expenditures | Pickle Lake, Nakina and Cochrane study areas | Energetic Balance Hypothesis: (focus: energy intake and expenditure component) | 1.1, 4.1 | Wiebe, P., CFS Thompson, I.D., CFS |
| 26 | 3.2.7 | Collaborative Woodland Caribou Research Program | Determining how biting fly abundance varies in response to disturbance and forest type | Nakina study areas | Energetic Balance Hypothesis: (focus: energy expenditure component) | 1.1, 4.1 | Raponi, M. Trent University Beresford, D., Trent University |
| 27 | 3.2.7 | Collaborative Woodland Caribou Research Program | Characterizing temporal changes in insect harassment and caribou behaviour in response to harassment | Pickle Lake, Nakina and Cochrane study areas | Energetic Balance Hypothesis: (focus: energy intake and expenditure component) | 1.1 | Rodgers, A.R., MNRF Shuter, J.L., MNRF Goren, A., MNRF Priadka, P., MNRF |
| 28 | 3.2.7 | Collaborative Woodland Caribou Research Program | Integrating results from caribou energy gains and losses into ecological energetics models for managed and unmanaged landscapes | Pickle Lake, Nakina and Cochrane study areas | Energetic Balance Hypothesis: (focus: synthesizing energy intake & energy expenditure results from other studies | 1.1, 4.1 | Thompson, I.D., CFS |
| 29 | 3.2.7 | Collaborative Woodland Caribou Research Program | Determining whether body condition and nutrition-related vital rates vary between managed and unmanaged landscapes | Pickle Lake, Nakina and Cochrane study areas | Energetic Balance Hypothesis- based predictions about differences in caribou condition and vital rates in managed vs. unmanaged landscapes | 1.1, 5.2 | Shuter, J.L., MNRF Rodgers, A.R., MNRF Brown, G.S., MNRF Thompson, I.T., CFS |
| 30 | 3.2.8 | Collaborative Woodland Caribou Research Program | Documenting traffic on the road networks in managed and unmanaged landscapes | Pickle Lake, Nakina and Cochrane study areas | Disturbance Hypothesis: (focus: characterizing disturbance patterns) | 3.73, 3.9, 7.2 | Hunt, L., MNRF |
| 31 | 3.2.8 | Collaborative Woodland Caribou Research Program | Determining whether caribou behaviour is affected by human road use | Pickle Lake | Disturbance Hypothesis: (focus: behavioural response to disturbance) | 3.73, 3.9, 7.2 | Shuter, J.L., MNRF Rodgers, A.R., MNRF Hunt, L., MNRF |
| 32 | 3.2.8, 3.2.10 | Collaborative Woodland Caribou Research Program | Evaluating the effectiveness of different approaches for restricting human road use | Pickle Lake, Nakina and Cochrane study areas | Multiple hypotheses about the factors that influence human compliance with road restrictions | 4.2.1, 5.5 | Hunt, L., MNRF |
| 33 | 3.2.8 3.2.10 | Collaborative Woodland Caribou Research Program | Evaluating the effectiveness of different road reclamation techniques | Several resource road locations throughout northwestern Ontario | Multiple hypotheses about the resources and conditions that might influence seedling establishment and re-growth on road beds | 4.2.1, 5.5 | Hall, J., MNRF Reid, D., MNRF |
| 34 | 3.2.9 | Monitoring and Assessment | Spatial comparison of occupancy modelling results – degree of overlap between predicted distributions of caribou, moose and wolves | Far North of Ontario | The Apparent Competition Hypothesis (focus: spatial overlap between general distribution of caribou, moose & wolves) | 5.2, 5.4, 5.5 | Poley, L.G., Trent University Pond, B.A., MNRF Schaefer, J.A., Trent University Brown, G.S., MNRF Ray, J.C., WCS Johnson, D.S., NOAA |
| 35 | 3.2.9 | Collaborative Woodland Caribou Research Program | Determining what resources and conditions influence wolf habitat selection at the landscape scale | Pickle Lake and Nakina study areas | All predation-related hypotheses (i.e. Apparent Competition, Predator Road Use, Prey Escape) | 5.2, 5.4, 5.5 | Kittle, A., Univ. of Guelph Patterson, B., MNRF Fryxell, J.M., Univ. of Guelph |
| 36 | 3.2.9, 3.2.10, 3.2.11 | Collaborative Woodland Caribou Research Program | Determining what resources and conditions influence habitat selection, territory size and density of different wolf packs | Pickle Lake, Nakina and Cochrane study areas | All predation-related hypotheses ( i.e. Apparent Competition, Predator Road Use, Prey Escape) | 5.2, 5.4, 5.5 | Anderson, M.L., Univ. of Guelph Kittle, A., Univ. of Guelph Patterson, B., MNRF Fryxell, J.M., Univ. of Guelph |
| 37 | 3.2.9 | Collaborative Woodland Caribou Research Program | Comparing within- day movements and activity levels among wolves, moose and caribou | Pickle Lake, Nakina and Cochrane study areas (wolf data) Dryden study area (historic moose data) | All predation-related hypotheses (i.e. Apparent Competition, Predator Road Use, Prey Escape) | 5.2, 5.4, 5.5 | Vander Vennen, L., Univ. of Guelph Patterson, B., MNRF Fryxell, J.M., Univ. of Guelph |
| 38 | 3.2.9 | Collaborative Woodland Caribou Research Program | Characterizing wolf diet composition in winter, using kill site investigation data | Pickle Lake, Nakina and Cochrane study areas (most data is from Nakina) | The Apparent Competition Hypothesis (focus: relative importance of caribou and alternate prey in wolf diets) | 5.2, 5.4, 5.5 | Moffatt, S., Univ. of Guelph Anderson, M.L. Vander Vennen, L., Univ. of Guelph Patterson, B., MNRF Fryxell, J.M., Univ. of Guelph |
| 39 | 3.2.9 | Collaborative Woodland Caribou Research Program | Characterizing wolf diet composition in winter - using stable isotope analysis | Pickle Lake, Nakina and Cochrane study areas | The Apparent Competition Hypothesis (focus: relative importance of caribou and alternate prey in wolf diets) | 5.2, 5.4, 5.5 | Patterson, B., MNRF |
| 40 | 3.2.9 | Collaborative Woodland Caribou Research Program | Characterizing wolf diet composition from spring-autumn - using scat content analysis | Nakina study area | The Apparent Competition Hypothesis (focus: relative importance of caribou and alternate prey in wolf diets) | 5.2, 5.4, 5.5 | Patterson, B., MNRF |
| 41 | 3.2.9 | Collaborative Woodland Caribou Research Program | Characterizing wolf diet composition from spring- autumn, using stable isotope analysis of hair samples | Pickle Lake, Nakina and Cochrane study areas | The Apparent Competition Hypothesis (focus: relative importance of caribou and alternate prey in wolf diets) | 5.2, 5.4, 5.5 | Patterson, B., MNRF |
| 42 | 3.2.9 | Collaborative Woodland Caribou Research Program | Characterizing bear diet composition during caribou calving season - using scat content analysis | Nakina study area | The Apparent Competition Hypothesis (focus: relative importance of caribou and alternate prey in bear diets) | 5.2, 5.4, 5.5 | Patterson, B., MNRF Rodgers, A.R., MNRF Shuter, J.L., MNRF Laporte, B., MNRF |
| 43 | 3.2.10 | Collaborative Woodland Caribou Research Program | Determining what factors influence wolf hunting efficiency | Nakina and Cochrane study areas | Two predation-related hypotheses (i.e. Apparent Competition & Predator Road Use) | 3.7.3, 3.9, 5.2, 5.5, 7.2 | Moffatt, S., Univ. of Guelph Patterson, B., MNRF Fryxell, J.M., Univ. of Guelph |
| 44 | 3.2.9 | Collaborative Woodland Caribou Research Program | Determining which landscape characteristics and conditions influence the probability that wolves will kill moose | Nakina study area | The Apparent Competition Hypothesis (focus: relative importance of caribou and alternate prey in wolf diets) | 5.2, 5.4, 5.5 | Kittle, A., Univ. of Guelph Patterson, B., MNRF Fryxell, J.M., Univ. of Guelph |
| 45 | 3.2.12, 3.2.3, 3.2.9, 3.2.10, 3.2.11 | Collaborative Woodland Caribou Research Program | Determining what general factors influence caribou survival | Pickle Lake, Nakina and Cochrane study areas | All major hypotheses assessed as part of the Collaborative Woodland Caribou Research Program (see Section 3.2.1) | 1.1, 5.2 | Shuter, J.L., MNRF Patterson, B., MNRF Rodgers, A.R., MNRF |
| 46 | 3.2.12 | Collaborative Woodland Caribou Research Program | Comparing condition at death of caribou living in managed and unmanaged forests | Pickle Lake, Nakina and Cochrane study areas | The Energetic Balance, Apparent Competition and Prey Escape Hypotheses | 1.1, 5.2 | Lowes, K., MNRF Rodgers, A.R., MNRF |
| 47 | 3.2.12 | Collaborative Woodland Caribou Research Program (offshoot) | Characterizing the diversity and prevalence of Protostrongylid parasites in caribou living in managed and unmanaged forests | Pickle Lake, Nakina and Cochrane study areas | Exposure to higher densities (and diversity) of alternate prey in managed landscapes will produce higher levels of parasite infection, poorer condition and/ or higher caribou mortality | 1.1, 5.2 | Verocai, G.G., Univ. of Calgary Kutz, S.J., Univ. of Calgary |
| 48 | 3.2.13, 3.2.14 | Collaborative Woodland Caribou Research Program | Develop and apply a spatial PVA model to assess caribou population viability and evaluate the potential consequences of different planning scenarios | Pickle Lake, Nakina and Cochrane study areas | All major hypotheses assessed as part of the Collaborative Woodland Caribou Research Program (see Section 3.2.1) | 3.7, 5.5, 7.2, 7.4 | Fryxell, J.M., Univ. of Guelph |
| 49 | 3.2.13, 3.2.14 | Collaborative Woodland Caribou Research Program | Apply a mechanistic model of caribou movement to determine how caribou respond to different resources and conditions | Pickle Lake, Nakina and Cochrane study areas | All major hypotheses assessed as part of the Collaborative Woodland Caribou Research Program (see Section 3.2.1) | 1.2, 3.7, 4.1, 4.2, 5.2, 5.4, 5.5, 7.2, 7.4 | Avgar, T., Univ. of Guelph Mallon, E., Univ. of Guelph Anderson, M.L., Univ. of Guelph Kittle, A.,Trent University Raponi, M. CFS Wiebe, P. MNRF |
| 50 | 3.2.14 | Spatial Ecology Program Caribou Resource Selection Function (RSF) Initiative | Generate caribou RSFs to help inform land use planning in the Far North of Ontario | The Continuous Distribution (4 Far North and 2 southern ranges: Spirit, Kinloch, Missisa, James Bay, Nipigon and Pagwachuan) | Several major hypotheses( see Section 3.2.14), which were adapted from those assessed as part of the Collaborative Woodland Caribou Research Program (see Section 3.2.1) | 3.1, 3.7, 3.9, 7.2, 7.4 | Hornseth, M., MNRF Rempel, R., MNRF |
Appendix 3-2: List of publications associated with caribou research projects in Ontario
Avgar, T., J. Baker, G.S. Brown, J. Hagens, E. Iwachewski, A. Kittle, E. Mallon, M. McGreer, A. Mosser, S.G. Newmaster, B. Patterson, D. Reid, A. Rodgers, J.L. Shuter, G. Street, I. Thompson, M. Turetsky, P. Wiebe, and J.M. Fryxell. (submitted manuscript). Space-use behaviour of woodland caribou based on a cognitive movement model
Avgar, T., A. Mosser, G.S. Brown and J.M. Fryxell. 2013. Environmental and individual drivers of animal movement patterns across a wide geographical gradient. Journal of Animal Ecology. 82(1): 96-106.
Avgar, T. 2013. From Diffusion to Cognition: Analytical, Statistical and Mechanistic Approaches to the Study of Animal Movement. Ph.D. Thesis. University of Guelph, Guelph, ON, Canada
Anderson, M.L. 2012. Wolf responses to spatial variation in moose density in northern Ontario. M.Sc.Thesis. University of Guelph, Guelph, ON, Canada
Kittle, A.M., B.R. Patterson, M.L. Anderson, S. Moffatt A.R. Rodgers, J.L. Shuter, D.E.B. Reid, J.A. Baker, G.S. Brown, I.D. Thompson, G.M. Street, T. Avgar, L. Vander Vennen, A. Mosser, J. Hagens, E. Iwachewski and J.M. Fryxell. (submitted manuscript). Wolves adapt territory size, not pack size to local habitat quality.
Kittle, A.M. (in prep.). The impact of prey, habitat and anthropogenic disturbance on space use by social carnivores. PhD thesis, University of Guelph. Mallon, E. 2014. Effects of Disturbance and Landscape Position on Vegetation Structure and Productivity in Ontario Boreal Forests: Implications for woodland caribou (Rangifer tarandus caribou) Forage. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada
Mosser, A., T. Avgar, G.S. Brown, C.S. Walker and J.M. Fryxell. 2014. Towards an energetic landscape: broad-scale accelerometry in woodland caribou. Journal of Animal Ecology. (Early View currently available online – DOI 10.1111/1365-2656.12187).
Kittle, A. (in prep.). Ph.D. Thesis. University of Guelph, Guelph, ON, Canada Raponi, M. (in prep.). Role of biting flies as a source of harassment for woodland caribou in Ontario. M.Sc. Thesis. Trent University, Peterborough, ON, Canada.
3.2.4
| Project # | Section in this report | Publications (to date) |
|---|---|---|
| 1 | 3.2.2 | Avgar, T., A. Mosser, G.S. Brown & J.M. Fryxell. 2013. Environmental and individual drivers of animal movement patterns across a wide geographical gradient. Journal of Animal Ecology. 82(1): 96-106. Avgar, T. 2013. From Diffusion to Cognition: Analytical, Statistical and Mechanistic Approaches to the Study of Animal Movement. Ph.D.Thesis. University of Guelph, Guelph, ON, Canada |
| 2 | 3.2.2 | Pond, B.A. Ecotype differentiation. pp. 43-49. in Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp. Wilson, K.S. 2013. Temporal and spatial variation in home range size for two woodland caribou ecotypes in Ontario. M.Sc. Thesis. Trent University, Peterborough, ON, Canada |
| 3 | 3.2.2 | Shuter, J.L., & Rodgers, A. R. 2012. Delineating demographic units of woodland caribou (Rangifer tarandus caribou) in Ontario: cautions and insights. Rangifer. 32(2), 159-181. |
| 5 | 3.2.2 | Thompson, L., and P.J. Wilson. Genetic structure. pp. 83-88 in Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp. Wilson, P.J. and L. Thompson. 2008. A preliminary genetic assessment of forest-dwelling and forest-tundra populations of woodland caribou in Ontario, Canada. Unpublished report to the Ontario Ministry of Natural Resources. May 2008. 10 pp. Thompson, L. (in prep.). Ph.D. Thesis. Trent University, Peterborough, ON, Canada Thompson, L., and P.J. Wilson. (in prep). Genetic Structure in Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Ministry of Natural Resources & Forestry. Klütsch, C.F.C., M. Manseau and PJ. Wilson. 2012. Phylogeographical Analysis of mtDNA Data Indicates Postglacial Expansion from Multiple Glacial Refugia in Woodland Caribou (Rangifer tarandus caribou). PLoS one 7(12): e52661. doi:10.1371/journal.pone.0052661 |
| 6 | 3.2.3 | Carr, N.L., A.R. Rodgers, S.R. Kingston, P.N. Hettinga, L.M. Thompson, J.L. Renton and P.J. Wilson, 2012. Comparative woodland caribou population surveys in Slate Islands Provincial Park, Ontario. Rangifer. 32(2), 205-218. |
| 7 | 3.2.3 3.2.4 | Poley, L.G., B.A. Pond, J.A. Schaefer, G.S. Brown, J.C. Ray and D.S. Johnson, 2013. Occupancy patterns of large mammals in the Far North of Ontario under imperfect detection and spatial autocorrelation. Journal of Biogeography. 1-11. Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp. |
| 11 | 3.2.4 | Avgar, T., A. Mosser, G.S. Brown & J.M. Fryxell. 2013. Environmental and individual drivers of animal movement patterns across a wide geographical gradient. Journal of Animal Ecology. 82(1): 96-106. Avgar, T. 2013. From Diffusion to Cognition: Analytical, Statistical and Mechanistic Approaches to the Study of Animal Movement. Ph.D. Thesis. University of Guelph, Guelph, ON, Canada McGreer, M., T. Avgar, J. Baker, G.S. Brown, J. Hagens, E. Iwachewski, A. Kittle, E. Mallon, A. Mosser, S.G. Newmaster, B. Patterson, D. Reid, A. Rodgers, J. Shuter, G. Street, I.D. Thompson, M. Turetsky, L.M. Vander Vennen, P. Wiebe and J.M. Fryxell. (submitted manuscript). Influence of spatial variation in forage availability and predation risk on habitat selection by woodland caribou (Rangifer tarandus caribou) in Ontario. McGreer, M. (in prep.). Influence of spatial variation in food availability and predation risk on habitat selection by woodland caribou in Ontario. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada Avgar, T., J. Baker, G.S. Brown, J. Hagens, E. Iwachewski, A. Kittle, E. Mallon, M. McGreer, A. Mosser, S.G. Newmaster, B. Patterson, D. Reid, A. Rodgers, J. Shuter, G. Street, I. Thompson, M. Turetsky, P. Wiebe, and J.M. Fryxell. (submitted manuscript). Space-use behaviour of woodland caribou based on a cognitive movement model. |
| 12 | 3.2.4 | Sherritt, A. (in prep.). Rapid assessment of woodland caribou habitat: the influence of habitat on site fidelity. M.Sc. Thesis. Trent University, Peterborough, ON, Canada |
| 13 | 3.2.4 | Wilson, K., G.S. Brown and B.A. Pond. Movement and activity analysis (home range and season delineation). pp. 50-54 in Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp. |
| 14 | 3.2.4 | Wilson, K.S. 2013. Temporal and spatial variation in home range size for two woodland caribou ecotypes in Ontario. M.Sc. Thesis. Trent University, Peterborough, ON, Canada Wilson, K., G.S. Brown and B.A. Pond. Movement and activity analysis (home range and season delineation). pp. 50-54 in Berglund, N.E., G.D. Racey, K.F. Abraham, G.S. Brown, B.A. Pond, and L.R. Walton. 2014. Woodland caribou (Rangifer tarandus caribou) in the Far North of Ontario: Background information in support of land use planning. Technical Report TR-147, Ministry of Natural Resources and Forestry, Thunder Bay, Ontario. 160 pp. |
| 15 | 3.2.5 | Webster, N.T.W. 2013. Forest management does not emulate natural disturbance with respect to plant diversity and forest community composition. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada |
| 16 | 3.2.5 | Kuzyk, R.E. 2013. Terrestrial lichen abundance in relation to stand structure and silvicultural history. M.Sc. Thesis. Lakehead University, Thunder Bay, ON, Canada |
| 17 | 3.2.5 | McMullin, R.T.; Thompson, I.D.; Newmaster, S.G.; Lacey, B.W. 2011. Estimating the biomass of woodland caribou forage lichens. Canadian Journal of Forest Research. 41: 1961-9. McMullin, R.T. 2011. Relationships Between Silviculture, Lichen Diversity and Woodland Caribou (Rangifer tarandus caribou) in Northern Ontario. Ph.D. Thesis. University of Guelph, Guelph, ON, Canada |
| 18 | 3.2.5 | Kuzyk, R.E. 2013. Terrestrial lichen abundance in relation to stand structure and silvicultural history. M.Sc. Thesis. Lakehead University, Thunder Bay, ON, Canada |
| 19 | 3.2.5 | McMullin, R.T.; F.W. Bell, S.G. Newmaster. 2012. The effects of triclopyr and glyphosate on lichens. Forest Ecology and Management. 264(15): 90-97 McMullin, R.T. 2011. Relationships Between Silviculture, Lichen Diversity and Woodland Caribou (Rangifer tarandus caribou) in Northern Ontario. Ph.D.Thesis. University of Guelph, Guelph, ON, Canada |
| 20 | 3.2.5 | McMullin, R.T.; I.D. Thompson, S.G. Newmaster. 2013. Lichen conservation in heavily managed boreal forests. Conservation Biology. 27(5): 1020-30. McMullin, R.T. 2011. Relationships Between Silviculture, Lichen Diversity and Woodland Caribou (Rangifer tarandus caribou) in Northern Ontario. Ph.D. Thesis. University of Guelph, Guelph, ON, Canada |
| 22 | 3.2.7 | Newmaster, S.G., R.A.D. Steeves, A.R. Rodgers, A.J. Fazekas, J.R. Maloles, R.T. McMullin and J.M. Fryxell. 2013. Examination of two new technologies to assess the diet of woodland caribou: video recorders attached to collars and DNA barcoding. Canadian Journal of Forest Research. 43:897-900. Thompson, I.D., M. Bakhtiari, A.R. Rodgers, J.A. Baker, J.M. Fryxell, E. Iwachewski. 2012. Application of a high-resolution animal-borne remote video camera with global positioning for wildlife study: Observations on the secret lives of woodland caribou. Wildlife Society Bulletin. 36(2):365-70. Thompson, I.D., P.A. Wiebe, E. Mallon, A.R. Rodgers, J.M. Fryxell, J.A. Baker. (submitted manuscript). Factors influencing the seasonal diet selection by woodland caribou in boreal forests in Ontario. |
| 23 | 3.2.7 | Mallon, E. 2014. Effects of Disturbance and Landscape Position on Vegetation Structure and Productivity in Ontario Boreal Forests: Implications for woodland caribou (Rangifer tarandus caribou) Forage. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada |
| 24 | 3.2.7 | Mosser, A., T. Avgar, G.S. Brown, C.S. Walker and J.M. Fryxell. 2014. Towards an energetic landscape: broad-scale accelerometry in woodland caribou. Journal of Animal Ecology. (Early View currently available online – DOI 10.1111/1365-2656.12187). |
| 26 | 3.2.7 | Raponi, M. (in prep.). Role of biting flies as a source of harassment for woodland caribou in Ontario. M.Sc. Thesis. Trent University, Peterborough, ON, Canada. |
| 34 | 3.2.9 | Poley, L.G., B.A. Pond, J.A. Schaefer, G.S. Brown, J.C. Ray and D.S. Johnson, 2013. Occupancy patterns of large mammals in the Far North of Ontario under imperfect detection and spatial autocorrelation. Journal of Biogeography. 1-11. |
| 35 | 3.2.9 3.2.10 3.2.11 | Kittle, A.M., B.R. Patterson, M.L. Anderson, S. Moffatt A.R. Rodgers, J.L. Shuter, D.E.B. Reid, J.A. Baker, G.S. Brown, I.D. Thompson, G.M. Street, T. Avgar, L. Vander Vennen, A. Mosser, J. Hagens, E Iwachewski and J.M. Fryxell. (submitted manuscript). Wolves adapt territory size, not pack size to local habitat quality. Kittle, A.M. (in prep.). The impact of prey, habitat and anthropogenic disturbance on space use by social carnivores. PhD thesis, University of Guelph. |
| 36 | 3.2.9 3.2.10 3.2.11 | Anderson, M.L. 2012. Wolf responses to spatial variation in moose density in northern Ontario. M.Sc.Thesis. University of Guelph, Guelph, ON, Canada Kittle, A.M., B.R. Patterson, M.L. Anderson, S. Moffatt A.R. Rodgers, J.L. Shuter, D.E.B. Reid, J.A. Baker, G.S. Brown, I.D. Thompson, G.M. Street, T. Avgar, L. Vander Vennen, A. Mosser, J. Hagens, E. Iwachewski and J.M. Fryxell. (submitted manuscript). Wolves adapt territory size, not pack size to local habitat quality. Kittle, A.M. (in prep.). The impact of prey, habitat and anthropogenic disturbance on space use by social carnivores. PhD thesis, University of Guelph. |
| 37 | 3.2.9 | Vander Vennen, L. (in prep.). Diel variability of wolf predation on ungulate prey. M.Sc. Thesis. University of Guelph, Guelph, ON, Canada |
| 38 | 3.2.9 | Moffatt, S. 2012. Time to event modelling: wolf search efficiency in northern Ontario. M.Sc.Thesis. University of Guelph, Guelph, ON, Canada |
| 43 | 3.2.10 | Moffatt, S. 2012. Time to event modelling: wolf search efficiency in northern Ontario. M.Sc.Thesis. University of Guelph, Guelph, ON, Canada |
| 46 | 3.2.12 | Lowes, K. 2013. Body condition of woodland caribou (Rangifer tarandus caribou) in managed and unmanaged Ontario forests. H.B.E.M.Thesis. Lakehead University, Thunder Bay, ON, Canada |
| 47 | 3.2.12 | Verocai, G. (in prep.). Ph.D. Thesis. University of Calgary, Calgary, AB, Canada |
| 49 | Avgar, T., R. Deardon and J.M. Fryxell. 2013. An empirically parameterized individual based model of animal movement, perception, and memory. Ecological Modelling. 251: 158-172. | |
| 50 | 3.2.14 | Hornseth, M.L. and R.S. Rempel. (submitted manuscript). Seasonal resource selection of woodland caribou (Rangifer tarandus caribou) habitat across a gradient of anthropogenic disturbance: A tool for land-use planning. |
Appendix 3-3: Population status and health measures: methodology, advantages & disadvantages
| Metric | Indicator type | Method | Advanatages | Disadvanatages |
|---|---|---|---|---|
| Adult survival rates (adult) | Population status |
|
|
|
| Pregnancy rates | Population status & caribou health |
|
|
|
| Live birth rates | Population status & caribou health |
|
|
|
| Calf survival rates (from birth to autumn) | Population status & caribou health |
|
|
|
| Recruitment rates (calf survival from birth to mid-winter) | Population status |
|
|
|
Appendix 3-4: General summary of research currently underway regarding caribou in Ontario
| Project # | General topic | Research topic | Specific research focus | Location | Hypothesis | CCP action item | Project leads (People) Organizational affiliation |
|---|---|---|---|---|---|---|---|
| 51 | Caribou population delineation | Monitoring and Assessment | Identify how disturbance affects functional landscape connectivity for caribou with respect to movement and gene flow, and whether thresholds or tipping points exists where changes in connectivity result in sub-structuring of populations. Determine how gene flow and population disturbance creates population structure structure of caribou at the northern range limit is affected by intermixing with the F-T ecotype. | Southern portion of the Continuous Distribution of Caribou, within the area of current commercial harvesting | The distribution of caribou in Ontario is spatially structured by the effects of disturbance (natural and anthropogenic) on landscape connectivity. Landscape fragmentation has different effects on male and female population genetic structure due to sex-biased dispersal. Where geographic locations for genetic exchange exist, the F-T ecotype has a genetic and demographic impact on sedentary caribou; alternatively, differences in behaviour and life-history events drive gene flow and population structure, effectively separating ecotypes genetically. | 1.3, 2.2, 7.4 | Viveen, A.,Trent University Brown, G.S., MNRF Wilson, P., Trent University |
| 52 | Caribou energetics | Collaborative Provincial Caribou Research Program | Developing a coarse resolution snow depth model and a mechanistic model of snow accumulation and melting | Pickle Lake, Nakina and Cochrane study areas | Caribou movements are impeded by snow conditions (e.g. depth, crusting) | 1.1, 4.1 | Wiebe, P., CFS Thompson, I.D., CFS |
| 53 | Caribou energetics and habitat | Collaborative Provincial Caribou Research Program | Determining what food selection strategy caribou use | Pickle Lake, Nakina and Cochrane study areas | Caribou food choice is selective Caribou food choice is opportunistic Caribou food choice is randomly | 1.1, 4.1 | Thompson, I. D., CFS |
| 54 | Caribou habitat | Collaborative Provincial Caribou Research Program | Evaluating seasonal habitat selection by caribou in response to the amount and arrangement of different forest types and natural and anthropogenic disturbances | Pickle Lake, Nakina and Cochrane study areas | Several alternative hypotheses about that factors that influence caribou use | 1.2, 4.1, 4.2 | Shuter, J.L. Rodgers, A.R. |
| 55 | Caribou habitat | Monitoring and Assessment | Evaluating longer- term effects of forest harvesting on caribou habitat conditions and behavioural responses. | Nakina and Cochrane study areas | Several alternative hypotheses about the short and long-term impacts of forest management activities on caribou | 1.2, 4.1, 4.2 | Donovan, V., Laurentian Univ. Brown, G.S., MNRF Mallory, F., Laurentian Univ. |
| 56 | Caribou habitat | Monitoring and Assessment | Using new geospatial data to develop methods that improve predictions of caribou habitat suitability and range distribution and design and test remote sensing-based indicators of caribou habitat suitability and quality | Areas throughout the Continuous Distribution (i.e. Far North and southern caribou ranges) | Use of new remote-sensed data sources (e.g. Lidar, RADAR sensor data) will improve efforts to identify and map caribou habitat | 1.2, 4.1, 4.2 | Hu, B., Univ. of Toronto Brown, G.S., MNRF Woods, M., MNRF Judah, A., Univ. of Toronto Zhang, W., Univ. of Toronto |
| 57 | Caribou habitat | Collaborative Provincial Caribou Research Program | Determining whether and how predation risk influences caribou movement behaviour and habitat selection | Nakina and Cochrane study areas | Increased levels of predation risk at the landscape scale, decreases the strength caribou responses to local scale predation risk | 1.2, 4.1, 4.2, 5.2, 5.4, 5.5 | McGreer, M., Guelph University M. Fryxell, J.M., Guelph University |
| 58 | Caribou re- occupancy or use of previously harvested stands | Collaborative Provincial Caribou Research Program | Determining whether previously harvested areas occupied by caribou have characteristics that distinguish them from other harvested areas | Nakina and Cochrane study areas | Caribou will inhabit and persist in previously harvested areas once suitable habitat conditions develop. | 1.0 | Reid, D., MNRF Shuter, J.L., MNRF Rodgers, A.R., MNRF |
| 59 | Road rehabilitation | Collaborative Provincial Caribou Research Program | Develop a new road classification system that reflects the road structural condition and road impacts on caribou, alternate prey and predators species | Nakina study area | Existing road classification systems are inadequate for characterizing current road conditions (e.g. level of re-vegetation, functional as animal movement corridors) and can be improved using remote- sensing-based methods. | 4.2.1, 5.5 | Stewart, J., Lakehead Univ. Reid, D., MNRF |
| 60 | Caribou mortality factors | Collaborative Woodland Caribou Research Program | Determining which landscapes characteristics influence predation-related mortality sites | Pickle Lake, Nakina and Cochrane study areas | All predation-related hypotheses (Apparent Competition, Predator Road Use, Prey Escape) | 5.2, 5.4, 5.5 | Shuter, J.L., MNRF Rodgers, A.R., MNRF Patterson, B., MNRF |
| 61 | Caribou mortality factors | Collaborative Woodland Caribou Research Program | Determining which landscapes characteristics influence caribou survival | Pickle Lake, Nakina and Cochrane study areas | All major hypotheses assessed as part of the Collaborative Woodland Caribou Research Program (see Section 3.2.1) | 1.1, 5.2 | Shuter, J.L., MNRF Patterson, B., MNRF Rodgers, A.R., MNRF |
Appendix 3-5- Overview of spatial population viability analysis (PVA) model for caribou: Structure and data used for initialization
Landscape attributes
- Model component
- Key attributes found to influence local movements and mortality rates of caribou were directly included in the model
- Data used for initialization
- GIS landscape variables including Ontario Land cover data, NDVI, elevation, linear features, fire occurrence, and snow cover
- These landscape variables were statistically linked to key biological attributes that influence caribou movements: land cover type, forage abundance, energetic gain, predation risk, insect harassment levels, and snow depth
- Time of day and year were also included
Caribou movement model [Project # 49. Please see Appendix 3-1]
- Model component
- Movement is modeled in 5 hour time steps
- Real caribou movement data are used to generate distributions of step lengths and turning angles (incorporated into a biased correlated random walk-based model of caribou movement
- Modeled caribou were more likely to move towards favourable habitat combinations and more likely to move away from unfavourable habitat combinations
- Data used for initialization
- Background caribou survival rates were estimated using data on fates of collared caribou.
- Landscape data used for habitat-specific predation-related mortality rates included Ontario Land cover data, moose abundance, moose resource selection function, wolf territory size and pack size in relation to prey abundance, NDVI, elevation, linear features, and snow cover data
Probability of death from predation
- Model component
- Modeled caribou interacted with predators and alternate prey and their probabilities of mortality were affected by these interactions
- Probability of predation varied throughout the landscape and depended on associated habitat attributes
- Data used for initialization
- Background and predation-related mortality rates were estimated using data on fates of collared caribou
- GIS landscape data used for habitat-specific predation-related mortality rates included Ontario Land cover data, NDVI, elevation, linear features, fire occurrence, and snow cover data
Caribou recruitment rates
- Model component
- Recruitment rates were influenced by the net energetic gain experienced by each caribou over the course of the previous year’s movements
- Approach used to model daily calf mortality risk incorporated demographic stochasticity (i.e. random variation in vital rates), which can have an important influence on population dynamics when population sizes are low
- Data used for initialization
- Background calf recruitment rates were estimated from the results of winter aerial surveys of the number of calves with collared caribou
Footnotes
- footnote[1] Back to paragraph Woodland caribou and reindeer are classified as belonging to the same species